
Name: Tyrannosaurus rex („König der Tyrannenechsen“)
Beschrieben: 1905 von Henry F. Osborn
Ordnung: Saurischia; Familie: Tyrannosauridae
Länge: ♂ bis zu 12m, ♀ bis 12,8m; Ausnahme-Exemplare über 13m
Gewicht: ♂ bis zu 7000kg, ♀ bis zu 8800kg; Ausnahmeexemplare über 12.000kg
Ernährung: carnivor
Beschreibung:
Tyrannosaurus ist der wohl berühmteste Dinosaurier der Welt und einer der größten Beutegreifer aller Zeiten. Er hat eine grünbraune, ledrige Haut, die von schwarzbraunen Streifen unterbrochen wird, wie bei einem Tiger. Die Männchen tragen darüber hinaus an den winzigen, aber sehr kräftigen Armen und im Nackenbereich tiefschwarze, dünne Filamentfedern. Diese dienen ihnen vor allem zur Arterkennung und während der Balz. Das dichte Federkleid, dass die Jungtiere noch tragen, verlieren sie im Laufe der Zeit. Junge Tyrannosaurier erreichen im Alter von drei Jahren etwa 400 – 500kg Gewicht. Die nächsten acht Jahre wachsen sie nur noch sehr langsam, bis sie als Teenager schließlich in einen extrem schnellen Wachstumsschub kommen. In der Pubertät nehmen sie jährlich fast eine Tonne an Gewicht zu. Mit 16 bis 18 Jahren werden sie geschlechtsreif. Ihr Wachstum verlangsamt sich danach wieder. Ein Tyrannosaurus kann bis zu 30 Jahre alt werden.

Jugendliche und Erwachsene Exemplare bevorzugen ganz unterschiedliche Beute. Hin und wieder schließen sie sich jedoch auch zusammen, um größere, in Herden lebende Beutetiere gemeinsam zur Strecke zu bringen. Die schnellen Jungtiere, die über 50km/h schnell laufen können, hetzen dabei ihre Beute und treiben sie vor sich her. Die älteren, langsameren, aber ungeheuer starken Erwachsenen, liegen dann schon im Unterholz auf der Lauer. Mit einem Überraschungsangriff bringen die Alten die Beute dann zu Fall und töten sie mit einem einzigen schnellen und ungeheuer kräftigen Biss. Zu seiner bevorzugten Beute zählen große Pflanzenfresser, wie Edmontosaurus und Triceratops.

Lebensweise:
Tyrannosaurus lebt in kleinen, lose organisierten Rudeln, die aus einem erwachsenen Weibchen, ihrem Partner und ihrem noch nicht geschlechtsreifen Nachwuchs bestehen. In einer T. rex-Familie geht es aber nicht gerade harmonisch zu. Meistens ziehen die Familienmitglieder allein durch die Wildnis und sammeln sich bloß dann, wenn sie es auf größere Beute abgesehen haben. Und um die gibt es oft Zoff. Streitigkeiten legt ein Tyrannosaurus fast immer mit brutaler Gewalt bei. Bisse, Tritte und Rammstöße mit dem Kopf werden zwar (zumeist!) nicht unbedingt mit tödlichen Absichten eingesetzt. Sie können allerdings schwere Verletzungen verursachen, so schwer, dass kaum ein Tyrannosaurus erwachsen wird, ohne dass sein Körper von tiefen Narben gezeichnet ist, die teils buchstäblich bis auf die Knochen reichen. Derartige Verletzungen stammen fast ausnahmslos von Artgenossen, sowohl aus der eigenen Familie als auch von rivalisierenden Banden, mit denen sie sich noch brutalere Kämpfe liefern.

Da die älteren Tiere jüngeren, gerade erst geschlechtsreif gewordenen in ihrer Physis massiv überlegen sind, kann ein ausgewachsenes Tyrannosaurus-Paar, dass sich ein Revier erobert hat, dieses meist über viele Jahre verteidigen und wird auch nur selten von Artgenossen herausgefordert. Stirbt eines der dominanten Tiere jedoch, reißen die sozialen Bande innerhalb des Rudels. Sogar die einander zuvor eng verbundenen Partner kennen keine Scheu, sich am Kadaver ihres einstigen Gefährten zu laben. Trotzdem sucht sich ein verwitweter Tyrannosaurus niemals wieder einen neuen Partner.

Das freigewordene Revier wird schnell durch ein jüngeres, meist gerade erst geschlechtsreif gewordenes Männchen besetzt, das sich dann schnell auf Brautschau begibt. Mit laut schallenden, weit hörbaren Tönen versucht der neue Revierbesitzer dann, ein Weibchen anzulocken. Nähert sich eine paarungsbereite Tyrannosaurus-Dame, legt er einen bizarren Paarungstanz an den Tag, wobei es seinen Nackenfederkamm aufstellt, seine Arme abspreizt, immer wieder kräftig mit den Füßen aufstampft und seine dann oft in einem hellen gelb gefärbte Kehle präsentiert.

Tyrannosaurus in Die Weißen Steine:
 |
Band I:Tyrannosaurus ist in diesem Buch der tierische Haupt-Antagonist und tritt in den Kapiteln „Das Monster im Nebel“ und „Joaquin“ in Erscheinung. Der merkwürdige Hügel, den Moritz in „Aliens und Zeitreisen“ entdeckt, ist ein Nisthaufen dieses Dinosauriers. Das aggresive Verhalten der Tiere dient jedoch lediglich zum Schutz ihres Nachwuchses. Sie sind eigentlich nicht besonders an Menschen als Beute interessiert. |
 |
Band II:Tyrannosaurus spielt in diesem Buch nur eine geringere Rolle. Das Brutpaar hat sich nach der wilden und traumatisierenden Jagd im Prolog in sein Revier zurückgezogen, um seine Jungen aufzuziehen. Im Kapitel „Der Bergpass“ tritt die Familie allerdings einmal in Erscheinung, ohne sich dabei aggressiv zu verhalten. |
 |
Band III:Moritz und seine Freunde laufen einem Tyrannosaurus direkt in die Arme, als sie in „Geburtstag“ nach der vermissten Sophie suchen. Das Buch endet hier für die Gruppe am See in einem Cliffhanger. Im Kapitel „Assisi“ entdecken Alicia und ihre Freundinnen in weiter Ferne ebenfalls einen einsamen Tyrannosaurus. |
 |
Band IV:Tyrannosaurus hat in diesem Buch die bisher meisten Auftritte und ist hier wieder der tierische Haupt-Antagonist, wobei aber kein Mensch tatsächlich durch ihn zu Schaden kommt. In den meisten Szenen stellt er keine Bedrohung dar, sodass man vor allem Einblicke in sein Verhalten bekommt. |
- Ein noch nicht ganz ausgewachsener Tyrannosaurus verfolgt Moritz und seine Freunde in „Der sterbende See“ bis zu ihrer Höhle, wo er sie tageland belagert. Moritz tauft das Exemplar auf den Namen „Narbenauge“.
- In „Erinnerungen“ werden Leon, Marie, Wasi, Hannah und John Zeuge einer dramatischen Jagdszene. Die T. rex-Familie erlegt dabei geneimsam einen jungen Triceratops.
- In „Asche und Blut“ begegnen Moritz und seine Freunde auf der Flucht vor dem Vulkan zunächst wieder Narbenauge und seinen Geschwistern, die sich aber ebenfalls nur auf der Flucht befinden und die Menschen nicht angreifen. Anders jedoch das männliche, ausgewachsene Tier: Es bewacht energisch seine Partnerin und treibt die Gruppe auseinander. Moritz und Melina werden in die eine, Max in eine andere und der Rest der Gruppe wieder in eine andere Richtung verschlagen.
- Ein Tyrannosaurus-Jungtier wandert in „Skylla und Charybdis“ am Strand entlang und trifft dabei auf John. Der macht auf das Tier mit seiner Fackel so einen Eindruck, dass es nicht wagt, ihn anzugreifen und einfach weiterzieht.
- Max wird in „Der lange Weg nach Hause“ von den jungen Tyrannosauriern gerettet: die halbwüchsigen Dakotaraptoren haben ihn bereits zu Boden gebracht, als die Rex-Jungtiere auftauchen. Sie töten einen der Dakotaraptoren und vertreiben den Rest.
- In „Drachen“ begegnen Moritz und Melina dem männlichen Tyrannosaurus erneut, der eine Gruppe Palaeosaniwa von dem Kadaver seiner Partnerin vertreibt. Doch ist das keineswegs als postume Zuneigung zu verstehen: der Tyrannosaurus frisst an dem Kadaver schließlich selbst.
 |
Hier geht’s zurück zur Hauptseite Leben in der Urzeit! |

Wissenschaftliches zu Tyrannosaurus:
 |
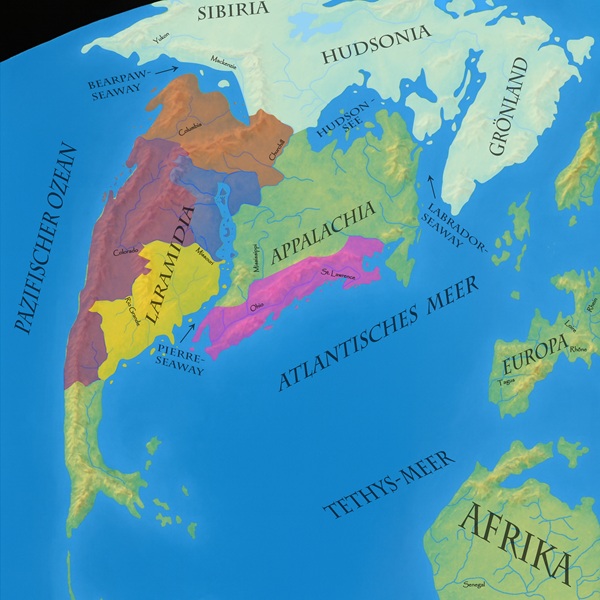
Tyrannosaurus rex – abgekürzt T. rex – war nicht nur der größte, sondern auch der am weitesten verbreitete Theropode des spätkreidezeitlichen Nordamerika. Seine Überreste wurden über den gesamten Westen des Kontinents hinweg gefunden. Vom Norden Kanadas bis weit nach Mexiko beherrschte T. rex seine Umwelt. Möglicherweise reichte sein Verbreitungsgebiet sogar noch bis nach Asien. Er ist in allen blauen, roten, violetten und gelben Gebieten sicher nachgewiesen und auch in nahezu allen Lagerstätten, die heute dort aufgeschlossen sind. Lediglich für eine Anwesenheit in den weit nördlich gelegenen Landschaften, hier weiß, fehlen Hinweise auf seine Existenz. |
Forschungsgeschichte
 Barnum Brown Barnum Brown |
Der berühmte Knochenjäger und Assistenzkurator am American Museum of Natural History Barnum Brown entdeckte im Jahr 1900 in der Lance Formation in Wyoming das erste teilweise erhaltene Skelett von Tyrannosaurus rex. |
Zwei Jahre später, im Jahr 1902, fand Brown ein weiteres, vollständigeres Teilskelett in einer heute als Hell Creek Formation bekannten Lagerstätte in Montana. Dieses bestand aus etwa 34 fossilen Knochen. Brown schrieb damals über diesen Fund:
„Steinbruch Nr. 1 enthält den Oberschenkelknochen, die Schambeine, den Oberarmknochen, drei Wirbel und zwei nicht näher bestimmte Knochen eines großen fleischfressenden Dinosauriers, der von Marsh noch nicht beschrieben wurde. … Ich habe im Kreidezeitalter noch nie etwas Vergleichbares gesehen.“1Dingus, L. & Norell, M. (2010). Barnum Brown – The man who discovered Tyrannosaurus rex. University of California Press, 90, 124.
 Henry F. Osborn Henry F. Osborn |
Im Jahr 1905 wies Henry F. Osborn, der damalige Präsident des American Museum of Natural History, dem zweiten Skelett die Inventarnummer AMNH 973 zu und gab ihm als neue Gattung und Art der Theropoden den Namen Tyrannosaurus rex. Der Gattungsname stammt aus dem Griechischen: τύραννος (tyrannos) bedeutet „Tyrann“, und σαῦρος (sauros) bedeutet „Echse“. Für den Artnamen verwendete Osborn das lateinische Wort rex, also „König“. Der vollständige wissenschaftliche Name lässt sich daher als „König der Tyrannenechsen“ oder „tyrannische Königsechse“ übersetzen.2Osborn, H. F. (1905). Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the AMNH, 21(14), 259–265. |
Um es vor Bombenangriffen infolge des zweiten Weltkriegs zu schützen, wurde dieses Skelett, der sogenannte Holotyp, 1941 an das Carnegie Museum of Natural History verkauft und nach Pittsburgh überführt, wo es sich noch heute befindet. Seither trägt es folglich eine andere Nummer: CM 9380. Der Preis betrug damals 7.000 Dollar. Inflationsbereinigt wären das heute mehr als 150.000 Dollar,3Dingus, L. & Norell, M. (2010). Barnum Brown – The man who discovered Tyrannosaurus rex. University of California Press, 90, 124. was aber im Vergleich zu den Preisen, zu denen Tyrannosaurus-Originalexemplare heutzutage gehandelt werden, immer noch ein Schnäppchen ist.

Doppelte Taufe: Dynamosaurus imperiosus
Ein anderes Exemplar, ein Teilskelett mit gut erhaltenem Unterkiefer, beschrieb Osborn im selben Jahr und Paper unter dem Namen Dynamosaurus imperiosus unter der Inventarnummer AMNH 973. Bereits 1906 erkannte Osborn jedoch, dass beide Skelette zur selben Art gehörten, und entschied sich dafür, den Namen Tyrannosaurus zu verwenden, den er zuerst, eine Seite vorher als, beschrieben hatte.4Osborn, H. F. & Brown, B. (1906). Tyrannosaurus, Upper Cretaceous carnivorous dinosaur. Bulletin of the AMNH, 22(16), 281–296. Heute wird der Unterkiefer im Natural History Museum in London ausgestellt, unter der Inventarnummer NHMUK R7994.5Breithaupt, B. H. et al. (2006). Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West. New Mexico Museum of Natural History and Science Bulletin, 35, 258.

Frühere Funde
 Arthur Lakes Arthur Lakes |
Doch eigentlich waren diese Exemplare gar nicht die ersten Spuren von Tyrannosaurus in der Forschungsgeschichte! Bereits 1874 entdeckte ein Student namens Peter T. Dotson auf dem South Table Mountain in Colorado einen gewaltigen Zahn, bei einer Grabung unter der Leitung von Arthur Lakes. |
 John B. Hatcher John B. Hatcher |
In den frühen 1890er-Jahren sammelte auch der Knochenjäger John B. Hatcher Knochenreste im östlichen Wyoming, die heute zu Tyrannosaurus gerechnet werden. Diese Fossilien wurden zunächst aber einem riesigen Ornithomiosaurier, benannt als „Ornithomimus grandis“, zugeschrieben. 6Breithaupt, B. H. et al. (2005). In celebration of 100 years of Tyrannosaurus rex – Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the earliest discoveries of Tyrannosaurus rex in the West. Abstracts with Programs, 37(7), 406. |
 Edward D. Cope Edward D. Cope |
1892 fand Edward D. Cope, einer der beiden Kontrahenten in den berüchtigten Bone Wars, zwei Wirbelfragmente eines großen Dinosauriers. Cope nahm an, dass diese Fragmente zu einem „agathaumiden“ Dinosaurier gehörten, also zu einem Vertreter der Ceratopsier. Er gab ihm den Namen Manospondylus gigas, was „riesiger poröser Wirbel“ bedeutet. |
Der Name bezog sich auf die zahlreichen Öffnungen für Blutgefäße, die er im Knochen erkannte.7Breithaupt, B. H. et al. (2005). In celebration of 100 years of Tyrannosaurus rex – Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the earliest discoveries of Tyrannosaurus rex in the West. Abstracts with Programs, 37(7), 406. Die Überreste von M. gigas wurden jedoch 1907 von Hatcher neu untersucht und als Knochen eines Theropoden und nicht eines Ceratopsiers identifiziert.8Hatcher, J. B. (1907). The Ceratopsia. Monographs of the United States Geological Survey, 49, 113–114.

Manospondylus gigas – Der Name Tyrannosaurus rex in Gefahr?
Henry F. Osborn hatte bereits 1917 die Ähnlichkeit zwischen Manospondylus gigas und Tyrannosaurus rex erkannt und darüber geschrieben. Zu diesem Zeitpunkt war allerdings bereits der zweite „poröse Wirbel“ verloren gegangen. Aufgrund des fragmentarischen Zustands der übrigen Wirbel setzte Osborn die beiden Gattungen nicht offiziell gleich, sondern betrachtete die ältere Gattung als unbestimmt.9Osborn, H. F. (1917). Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bulletin of the American Museum of Natural History, 35(43), 733–771.
 Peter Larson Peter Larson |
Im Juni 2000 entdeckte das Black Hills Institute unter der Leitung von Peter Larson etwa zehn Prozent eines Tyrannosaurus-Skeletts an einer Fundstelle, die sehr wahrscheinlich der ursprünglichen Fundstelle von M. gigas entspricht. |
Nach diesem Fund hätte der Name Tyrannosaurus rex eigentlich gemäß der Prioritätsregel des International Code of Zoological Nomenclature (ICZN) aufgegeben werden müssen, was Larson auch vorschlug, nachdem er dieses Exemplar für dasselbe Tier wie den Holotyp von M. gigas auswies.10Larson, N. L. (2008). One hundred years of Tyrannosaurus rex – the skeletons. In Larson, P. & Carpenter, K. (Hgg.), Tyrannosaurus rex, the Tyrant King. Indiana University Press, 1–55.
 Christopher Brochu Christopher Brochu |
Andere Forscher, darunter Christopher Brochu, waren diesbezüglich aber skeptisch. Er argumentierte, dass beide Taxa zwar wahrscheinlich dieselbe Art darstellen, es jedoch unmöglich sei zu beweisen, dass das vom Black Hills Institute entdeckte Exemplar tatsächlich genau dasselbe Tier ist wie M. gigas, da der Holotyp von M. gigas sehr schlecht erhalten ist. |
Zudem besagt eine Änderung des Internationalen Codes der Zoologischen Nomenklatur aus dem Jahr 2000, dass ein in der Öffentlichkeit weit verbreiteter Name wie T. rex Vorrang vor einem Namen mit rein technischer Priorität hat, wenn dieser seit dem Jahr 1899 über mehr als fünfzig Jahre hinweg nicht mehr als gültiger Name in der wissenschaftlichen Literatur verwendet wurde, wie es bei M. gigas der Fall ist. Tyrannosaurus rex darf seinen ikonischen Namen also behalten.11Brochu, C. R. (2003). Osteology of Tyrannosaurus rex – insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Society of Vertebrate Paleontology Memoirs, 7, 1–138. doi:10.2307/3889334
Neuere Forschungen
Von den 1910er-Jahren bis zum Ende der 1950er-Jahre blieben die Funde von Barnum Brown die einzigen bekannten Exemplare von T. rex. Die Weltwirtschaftskrise und mehrere Kriege hielten viele Paläontologen davon ab, weitere Feldforschungen in den Lagerstätten des mittleren Westens durchzuführen. Erst seit den 1960er-Jahren nahm das wissenschaftliche Interesse an T. rex wieder stark zu. In dieser Zeit wurden in West-Nordamerika insgesamt 42 Skelette entdeckt, deren Erhaltungszustand zwischen etwa fünf und achtzig Prozent lag. Besonders viele neue Funde wurden in den 1990er-Jahren gemacht, fast doppelt so viele wie in allen vorherigen Jahrzehnten zusammen.12Larson, N. L. (2008). One hundred years of Tyrannosaurus rex – the skeletons. In Larson, P. & Carpenter, K. (Hgg.), Tyrannosaurus rex, the Tyrant King. Indiana University Press, 1–55. Dazu gehörten auch einige der vollständigsten und berühmtesten Tyrannosaurus-Skelette überhaupt. An diesen habe ich mich bei der Rekonstruktion „meiner“ Tyrannosaurier in der Romanreihe orientiert:
„Sue“ (FMNH PR2081)
|
Das T. rex-Weibchen in meinem Roman habe ich dem Exemplar „Sue“ nachempfunden. Dieses verdankt seinen Namen der Amateurpaläontologin Sue Hendrickson, die am 12. August 1990 in der Hell Creek Formation nahe Faith in South Dakota ein etwa zu 90 Prozent erhaltenes T. rex-Skelett entdeckte. Es war das bis dahin vollständigste und größte überhaupt bekannte und wurde deshalb zu Ehren seiner Entdeckerin benannt. Es ist aber – und das gilt auch für alle anderen der hier genannten Exemplare – nicht bekannt, ob „Sue“ wirklich ein Weibchen oder doch ein Männchen war! |

Kurs nach „Sues“ Entdeckung und Präparation durch das Black Hills Institute in Hill City in South Dakota entbrannte jedoch ein unangenehmer Rechtsstreit um die Besitzverhältnisse des spektakulären Fossils. Maurice Williams, der Eigentümer des Grundstücks, auf dem es gefunden worden war, sowie der Sioux-Stamm, dem er angehörte, beanspruchten das Exemplar für sich. Das Black Hills Institute war dagegen davon ausgegangen, es rechtmäßig erworben zu haben, allerdings für einen geradezu lächerlich günstig erscheinenden Preis von nur 5.000 Dollar. 1992 führte das FBI im Rahmen der Ermittlungen eine Razzia im Black Hills Institute durch und beschlagnahmte dabei das Exemplar. 1997 wurde der Rechtsstreit zugunsten von Williams entschieden, und das Fossil ging in dessen Besitz über. Williams ließ „Sue“ daraufhin über das Auktionshaus Sotheby’s in New York versteigern. Das Field Museum of Natural History in Chicago erwarb das Fossil schließlich für 8,4 Millionen US-Dollar.13Fiffer, S. (2000). Tyrannosaurus Sue. W. H. Freeman and Company, New York. 195-212.14Escobedo, T. (2014). When the feds seized a T. rex. CNN.15Browne, M. (1996). Fossil Dealer, Target of Federal Prosecutors, Begins Jail Term. New York Times.16Regalado, A. (1995). Fossil dealers innocent – and guilty. Science, 267(5205). Dies war damals der höchste Preis, der jemals für ein Fossil gezahlt wurde, bis dieser Rekord später vom Tyrannosaurus-Exemplar „Stan“ übertroffen wurde.17Small, Z. (2020). T. rex Skeleton Brings $31.8 Million at Christie’s Auction. The New York Times. „Sue“ besitzt eine Länge von 12,3 bis 12,4 Metern und erreichte an der Hüfte eine Höhe von etwa 3,66 bis 3,96 Metern. Das Lebendgewicht wird auf etwa 8,4 Tonnen geschätzt.18Hutchinson, J. R. et al. (2011). A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth. PLOS ONE, 6(10), e26037. doi:10.1371/journal.pone.0026037
„Stan“ (NHMAD 2020.00001)
|
Ein weiteres berühmtes Exemplar, dass in meinem Roman Pate für das T. rex-Männchen stand, wurde nach dem Amateurpaläontologen Stan Sacrison benannt. Es wurde 1987 ebenfalls in der Hell Creek Formation gefunden, nahe Buffalo in South Dakota und ist das zweitvollständigste bekannte T. rex-Skelett. 199 Knochen wurden damals geborgen, etwa siebzig Prozent des gesamten Skeletts. Stan gehört damit zu den bekanntesten und vollständigsten Tyrannosaurus rex-Funden. |
.jpg)
„Stan“ diente als Vorlage für jede Menge Abgüsse, die nun in Museen auf der ganzen Welt ausgestellt sind. Das Original ist seit 2025 im neuen Naturkundemuseum von Abu Dhabi zu sehen, nachdem er für 31,8 Milliomen Dollar ersteigert wurde.19Small, Z. (2020). T. rex Skeleton Brings $31.8 Million at Christie’s Auction. The New York Times.20Greshko, M. (2022). Stan the T. rex found! World’s most expensive fossil finds home in a new museum. National Geographic.21Martin, N. (2025). Exclusive. New Natural History Museum Abu Dhabi features two fighting T. rexes. National Geographic Society. Dort erhielt „Stan“ die neue Inventarnummer NHMAD 2020.00001.22Boeye, A. T. et al. (2026). Evidence of bird-like foot function in Tyrannosaurus. Royal Society Open Science, 13(2). doi:10.1098/rsos.252139 „Stan“ ist insgesamt 11.78 Meter lang. Sein Körper zeigt zahlreiche Knochenverletzungen, darunter gebrochene und wieder verheilte Rippen, einen gebrochenen Halswirbel sowie ein etwa 2,5 Zentimeter breites Loch im Hinterkopf, das wahrscheinlich durch den Biss eines anderen Tyrannosaurus verursacht wurde. Zwei Halswirbel sind miteinander verwachsen, und ein weiterer Wirbel zeigt eine aauffällige Knochenwulst. Diese Verletzungen könnten ebenfalls durch einen Biss eines anderen Tyrannosaurus entstanden sein. Da alle Verletzungen verheilt waren, steht fest, dass Stan diese Angriffe überlebt hatte.23Tanke, D. H. & Currie, P. J. (1998). Head-biting behavior in theropod dinosaurs. Paleopathological evidence. Gaia, 15, 167–184.
„Bucky“ (TCM 2001.90.1)
|
1998 entdeckte der damals erst zwanzigjährige Rancher und Cowboy Bucky Derflinger einen aus dem Boden ragenden Zeh eines T. rex nahe der Stadt Faith in South Dakota. Damit wurde er zum jüngsten Menschen, der je ein Exemplar dieser Art entdeckt hat. Das Fossil erhielt ebenfalls den Namen des Entdeckers. 24Larson, P. L. & Carpenter, K. (2008). Tyrannosaurus rex, the Tyrant King. Life of the Past. Indiana University Press. Von „Bucky“ habe ich mich zu meinem Antagonisten „Narbenauge“ in meinem Buch inspirieren lassen. |

Bucky war außergewöhnlich gut erhalten und konnte vom Black Hills Institute in South Dakota vergleichsweise leicht präpariert werden. Heute befindet sich das Exemplar mit der Inventarnummer TCM 2001.90.1 im The Children’s Museum of Indianapolis in Indiana und wird gemeinsam mit einem Abguss des erwachsenen Tyrannosaurus-Exemplar „Stan“ in einer Jagdszene präsentiert. Beide Tyrannosaurus-Individuen greifen dabei einen Triceratops namens „Kelsey“ an. Er ist der erste noch nicht ausgewachsene Tyrannosaurus, der dauerhaft in einem Museum ausgestellt wurde.25Crosslin, R., Fortney, M. & Dinosphere Exhibit Development Team (2004). Resource Materials. Bucky-Tyrannosaurus rex. Dinosphere: A 3–5 Unit of Study. Außerdem ist „Bucky“ das erste T. rex-Skelett, bei dem eine Furcula, also ein Gabelbein oder Wunschbein, erhalten ist. Außerdem besitzt Bucky einen nahezu vollständigen Satz von Gastralia (Bauchrippen) sowie eine Elle (Ulna), also einen der beiden Unterarmknochen.26Larson, P. The First Recognized Furcula for Tyrannosaurus rex. Black Hills Institute of Geological Research. Er ist etwa drei Meter hoch und elf Meter lang. Nach dem Tod des Tieres wurden die Knochen durch Wasser verfrachtet und lagerten sich schließlich in einer flachen Senke gemeinsam mit Knochen von Edmontosaurus und Triceratops ab. Bis heute konnten 101 Knochen identifiziert und wissenschaftlich bestätigt werden, das entspricht etwa 34 % des gesamten Skeletts.
„Jane“ (BMRP 2002.4.1)
|
Im Sommer 2001 entdeckten Carol Tuck und Bill Harrison bei einer Expedition des Burpee Museum of Natural History in Rockford im Bundestaat Illinois ein etwa fünfzig Prozent vollständiges Skelett in der Hell Creek Formation. Dieses Fossil erhielt den Spitznamen „Jane“, der zu Ehren von Jane Solem, einer Förderin des Museums, vergeben wurde. Als ich die „Halbstarken“, also die drei Jungtiere des T. rex-Rudels in meinem Roman entworfen habe, war „Jane“ dabei die wichtigste Quelle. |
Nach vier Jahren Präparationsarbeit wurde „Jane“ im als Hauptstück der Ausstellung Jane: Diary of a Dinosaur präsentiert. Sie war und ist bis heute Gegenstand einer wissenschaftlichen Kontroverse: die Mehrheit der Forscher hielt sie bis vor kurzem noch für einen jugendlichen T. rex, doch viele andere Experten interpretierten sie als Vertreterin einer eigenen, viel kleineren Tyrannosaurier-Art namens Nanotyrannus. Peter Larson stellte 2013 fest, dass Jane in mehreren Merkmalen stärker den Exemplaren CMNH 7541 und LACM 28471 ähnelt als erwachsenen Tyrannosaurus rex. Dazu gehören eine höhere Zahnzahl, ein großes pneumatisches Foramen im Quadratojugale, ein T-förmiger Postorbitalknochen, sowie verwachsene Schulterblätter und Beckenknochen.27Larson, P. L. (2013). The case for Nanotyrannus. In Parrish, J. M., Molnar, R. A., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. University of Indiana Press, 15–53. Paläontologen, die Jane als jugendlichen Tyrannosaurus rex betrachten, schätzen das Alter des Tieres zum Zeitpunkt seines Todes auf etwa elf Jahre. Das rekonstruierte Skelett erreicht eine Länge von ungefähr 6,5 Metern und ein Gewicht von etwa 954 Kilogramm.28Hutchinson, J. R. et al. (2011). A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth. PLOS ONE, 6(10), e26037. doi:10.1371/journal.pone.0026037
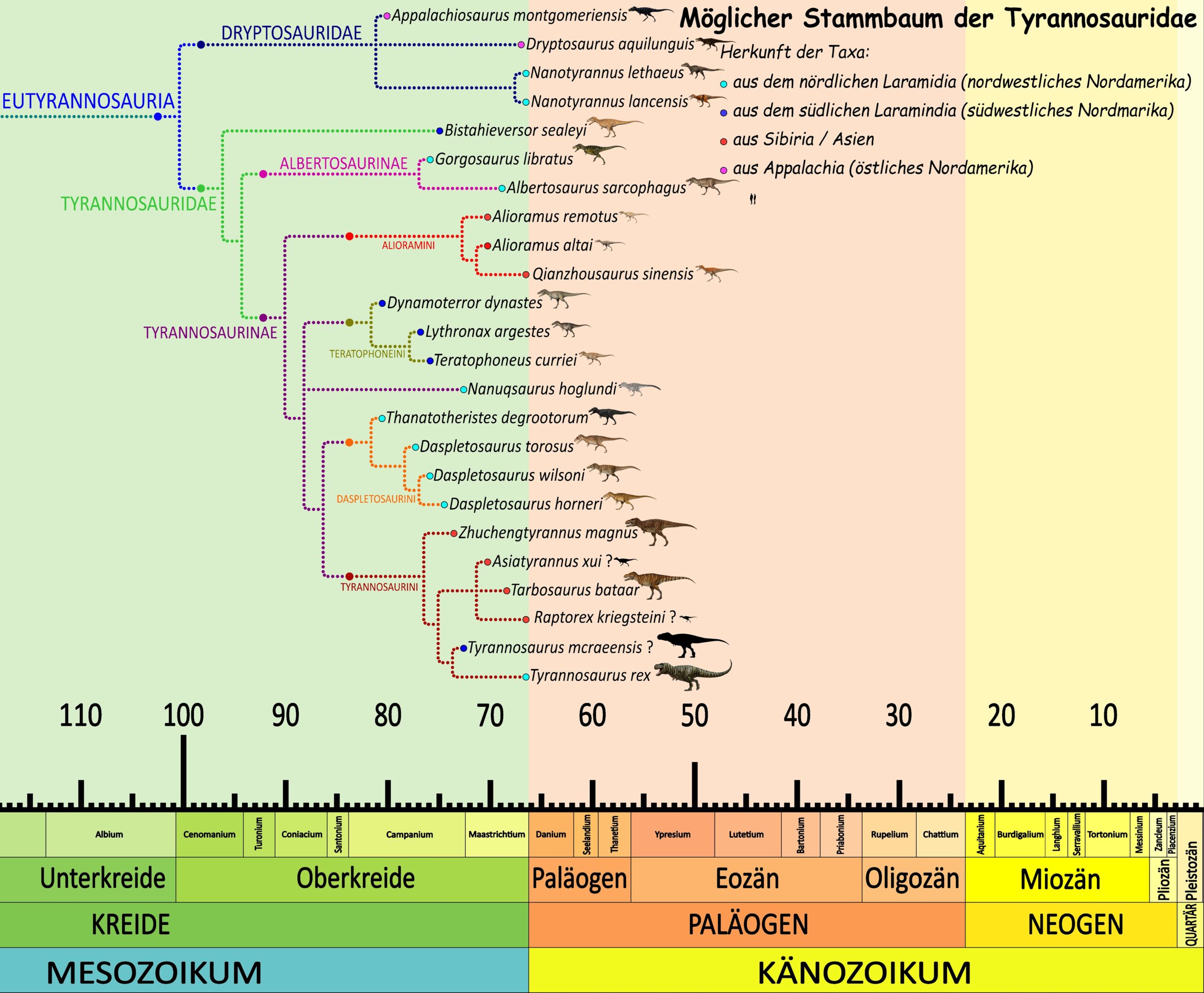
Lindsay Zanno und James G. Napoli vertraten 2025 die Auffassung, dass Tyrannosaurus und Nanotyrannus zwei eigenständige Gattungen seien. In diesem Zusammenhang ordneten sie „Jane“ einer eigenen Art von Nanotyrannus als Holotypus zu: Nanotyrannus lethaeus.29Zanno, L. E. & Napoli, J. G. (2025). Nanotyrannus and Tyrannosaurus coexisted at the close of the Cretaceous. Nature, 648(8093), 357–367. doi:10.1038/s41586-025-09801-6 Im Jahr 2026 führten Holly Woodward, Nathan Myhrvold und Jack Horner eine umfassende histologische Untersuchung von 17 Tyrannosaurier-Exemplaren durch. Dabei stellten sie fest, dass die Wachstumskurven von „Jane“ und einem weiteren jungen Exemplar (BMRP 2006.4.4) nicht gut zu den Wachstumsmustern anderer Tyrannosaurus-Exemplare passen. Dies würde zu der Nanotyrannus-Hypothese passen. Die Autoren betonen jedoch ausdrücklich, dass diese Abweichungen allein nicht ausreichen, um die Eigenständigkeit von Nanotyrannus als gültige Gattung zu beweisen.30Woodward, H. N., Myhrvold, N. P. & Horner, J. R. (2026). Prolonged growth and extended subadult development in the Tyrannosaurus rex species complex revealed by expanded histological sampling and statistical modeling. PeerJ, 14, e20469. doi:10.7717/peerj.20469 Ob Jane nun wirklich ein Tyrannosaurus oder doch ein Nanotyrannus ist, muss somit noch weiter erforscht werden.

Anhand dieser, aber auch vieler weiterer gefundener Exemplare haben Forscher auf der ganzen Welt inzwischen einige sehr erstaunliche Erkenntnisse gewonnen. Jedes Jahr erscheinen neue Studien zu seiner Anatomie, seiner Lebensweise und seines möglichen Verhaltens. Damit ist T. rex einer der am besten erforschten Dinosaurier überhaupt und derjenige Bewohner der Hell Creek Formation, bei dem ich mir die wenigsten literarischen Freiheiten erlaubt habe.
|
Bis heute wurde allerdings noch kein nur wenige Tage bzw. Wochen altes T. rex-Baby gefunden. Wir wissen also bislang noch nicht, wie die frischgeschlüpften Jungtiere ausgesehen haben. Die „fluffige“ Darstellung in meinem Buch ist hier also einmal tatsächlich reine Fiktion. |
Wachstum und „Lebensgeschichte“

Der Weg vom Schlüpfling zum ausgewachsenen Tier galt bei T. rex bis vor kurzem allerdings noch als sehr gut rekonstruiert. Histologische Analysen an Knochen verschiedener Exemplare hatten gezeigt, dass das Jugendwachstum einer S-förmigen Kurve folgte: Junge Tiere sollen bis etwa zum 14. Lebensjahr unter 1.800 kg geblieben sein, danach habe eine etwa vier Jahre dauernde Phase extrem schnellen Wachstums eingesetzt, in der sie durchschnittlich rund 600 kg pro Jahr zunahmen. Mit etwa 16 bis 18 Jahren soll sich das Wachstum deutlich verlangsamt haben, sodass sich selbst bei deutlich älteren Tieren nur noch geringe Gewichtsunterschiede feststellen lassen.37Horner, J. R. & Padian, K. (2004). Age and growth dynamics of Tyrannosaurus rex. Proceedings: Biological Sciences, 271(1551), 1875–1880. doi:10.1098/rspb.2004.2829 So betrug der Gewichtsunterschied zwischen der 28 Jahre alten Sue und einem 22 Jahre alten kanadischen Exemplar (RTMP 81.12.1) nur etwa 600 kg.38Erickson, G. M. et al. (2006). Tyrannosaur life tables – an example of nonavian dinosaur population biology. Science, 313(5784), 213–217. doi:10.1126/science.1125721 In der Phase des extremen Wachstumsschubs hingegen hätte ein T. rex dagegen bis zu etwa 1.790 kg pro Jahr zugenommen.39Hutchinson, J. R. et al. (2011). A computational analysis of limb and body dimensions in Tyrannosaurus rex with implications for locomotion, ontogeny, and growth. PLoS ONE, 6(10), e26037. doi:10.1371/journal.pone.0026037 Der abrupte Rückgang der Wachstumsrate nach der schnellen Wachstumsphase wiederum könnte ein Hinweis auf das Erreichen der Geschlechtsreife sein. Dies wird auch durch den Fund von medullärem Knochengewebe im Oberschenkelknochen von „B-Rex“, einem 16 bis 20 Jahre alten T. rex aus Montana gestützt. Solches Knochengewebe kommt bei heutigen Vögeln nur bei Weibchen während der Eiablage vor und zeigt daher an, dass dieses Tier geschlechtsreif gewesen sein muss.40Schweitzer, M. H. et al. (2005). Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. Science, 308(5727), 1456–1460. doi:10.1126/science.111215841Lee, A. H. & Werning, S. (2008). Sexual maturity in growing dinosaurs does not fit reptilian growth models. Proceedings of the National Academy of Sciences, 105(2), 582–587. doi:10.1073/pnas.070890310542Schweitzer, M. H. et al. (2016). Chemistry supports the identification of gender-specific reproductive tissue in Tyrannosaurus rex. Scientific Reports, 6, 23099. doi:10.1038/srep23099
Basierend auf diesen Studien habe ich „meinem“ T. rex in Die Weißen Steine einen entsprechenden Lebenszyklus gegeben. Auch das Familienleben habe ich entsprechend so dargestellt, dass die unterschiedlich großen Individuen unterschiedliche Rollen im Verhalten und auch im Ökosystem einnehmen.

Doch diese Darstellung könnte neueren Studien zufolge nicht mehr wahrscheinlich sein. Eine umfassende histologische Analyse von 17 Tyrannosaurier-Exemplaren konnte 2026 an den ältesten Exemplaren deutlich mehr „Wachstumsringe“ feststellen als bisher sichtbar. Die Autoren argumentierten daher, dass das Wachstum von T. rex wahrscheinlich deutlich langsamer und gleichmäßiger verlief als in früheren Studien angenommen. Demnach könnte es eine längere Phase subadulter Entwicklung gegeben haben, wobei die maximale Körpergröße erst im Alter von etwa 35 bis 40 Jahren erreicht wurde. Gleichzeitig stellten die Autoren fest, dass die Wachstumsmuster zweier unersuchter Exemplare, darunter „Jane“, nicht zu den anderen T. rex-Fossilien im Modell passten.43Woodward, H. N. et al. (2026). Prolonged growth and extended subadult development in the Tyrannosaurus rex species complex revealed by expanded histological sampling and statistical modeling. PeerJ, 14, e20469. doi:10.7717/peerj.20469 Somit könnte „Jane“ tatsächlich eine andere Spezies, also einen Nanotyrannus darstellen, wie auch kurz zuvor in einer Studie von Zanno und Napoli (2025) vorgeschlagen wurde.44Zanno, L. E. & Napoli, J. G. (2025). Nanotyrannus and Tyrannosaurus coexisted at the close of the Cretaceous. Nature. doi:10.1038/s41586-025-09801-6 Wenn aber „Jane“, die für viele der oben genannten Studien eines der wichtigsten Referenzexemplare war, aber nun gar kein T. rex ist, dann müssen sämtliche bisherige Datensätze zu den Wachstumsmodellen noch einmal überprüft werden. Und die These, dass T. rex schnell wuchs und jung starb,45Paul, G. S. (2008). Chapter 18 – The extreme life style and habits of the gigantic tyrannosaurid superpredators of the Cretaceous North America and Asia. In Larson, P. L. & Carpenter, K. (Hgg.), Tyrannosaurus, the Tyrant King. Indiana University Press, 307–345.46Holtz, T. R. Jr. (2013). The life and times of Tyrannosaurus rex, with Dr. Thomas Holtz. Burke Museum of Natural History and Culture. könnte dann nicht mehr so stehen bleiben. Hier gilt es nun, weitere Studien und Revisionen abzuwarten.
Körperhaltung und Funktion der Arme
Wie viele zweibeinige Dinosaurier wurde auch T. rex historisch lange als eine Art „lebendes Dreibein“ dargestellt: mit stark aufgerichtetem Körper und einem Schwanz, der über den Boden schleifte, ähnlich wie bei einem Känguru. Diese Vorstellung geht auf Joseph Leidys Rekonstruktion von Hadrosaurus aus dem Jahr 1865 zurück, die erste Darstellung eines Dinosauriers in aufrechter zweibeiniger Haltung.47Leidy, J. (1865). Memoir on the extinct reptiles of the Cretaceous formations of the United States. Cretaceous reptiles of the United States. Smithsonian Contributions to Knowledge, 14, 1–4. doi:10.5962/bhl.title.39830 1915 übernahm Henry F. Osborn dieses Bild auch für seinen T. rex, als er im American Museum of Natural History das erste Skelettmodell in dieser Pose aufbauen ließ. Für 77 Jahre blieb es so stehen, bis es 1992 umgebaut wurde. Die fehlerhafte Rekonstruktion des AMNH beeinflusste jedoch jahrzehntelang Filme und Illustrationen, darunter auch Klassiker wie Gwangis Rache (1969). Hierbei sei allerdings angemerkt, dass „Gwangi“ eigentlich ein Allosaurus sein soll. Offensichtlich hat sich der Stop-Motion-Künstler Ray Harryhausen aber dennoch von historischen Tyrannosaurus-Darstellungen wie von Charles R. Knight inspirieren lassen.
Spätestens in den 1970ern setze sich aber allmählich die Erkenntnis durch, dass die Känguru-Haltung biomechanisch nicht möglich war. Moderne Darstellungen in Museen, Kunst und Film zeigen T. rex heute mit nahezu waagerecht gehaltenem Körper und einem nach hinten ausgestreckten Schwanz, der den Kopf ausbalanciert. Zum Hinsetzen und Hinlegen könnte T. rex sein Gewicht auf eine verdickte Struktur des Schambeins gestützt haben. Um wieder aufzustehen, nutzte er zum Hochdrücken vielleicht auch seine kleinen, aber dennoch sehr kräftigen Arme.48Newman, B. H. (1970). Stance and gait in the flesh-eating Tyrannosaurus. Biological Journal of the Linnean Society, 2(2), 119–123. doi:10.1111/j.1095-8312.1970.tb01707.x Doch ist diese These umstritten. Verheilten Ermüdungsbrüche in den Armknochen wurden debei unterschiedlich interpretiert: Einige Forscher sehen darin einen Hinweis darauf, dass die Arme kaum nützlich waren und wohl kaum für einen Push-Up gebraucht werden konnten.49Padian, K. (2022). Why tyrannosaurid forelimbs were so short. An integrative hypothesis. Acta Palaeontologica Polonica, 67(1), 63–76. Andere betrachten sie gerade als Beleg dafür, dass die Arme tatsächlich immer wieder in Gebrauch waren und wie der restliche Körper auch hin und wieder Verletzungen erlitten.50Stevens, K. A., Larson, P., Willis, E. D. & Anderson, A. (2008). Rex, sit. Digital modeling of Tyrannosaurus rex at rest. In Larson, P. & Carpenter, K. (Hgg.), Tyrannosaurus rex, the tyrant king. Indiana University Press, 192–203.
 Lawrence M. Lambe Lawrence M. Lambe |
Als T. rex erstmals entdeckt wurde, war der Aufbau der Arme übrigens noch gar nicht bekannt, nur der Oberarmknochen. Und so hatte T. rex in der ersten Rekonstruktion von 1915 noch weit längere, dreifingrige Vorderarme, wobei sich Osborn an denen von Allosaurus orientierte. Bereits ein Jahr vor der Ausstellungseröffnung hatte jedoch Lawrence M. Lambe die kurzen zweifingrigen Vorderarme des nah verwandten Gorgosaurus beschrieben. Dies deutete stark darauf hin, dass auch T. rex ähnliche Vordergliedmaßen besaß.51Lambe, L. M. (1914). On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. Ottawa Naturalist, 27, 129–135. |
Bestätigt wurde dies allerdings erst 1989, als die ersten vollständigen T. rex-Arme am Exemplar „Wankel Rex“ (MOR 555) entdeckt wurden.52Horner, J. R. & Lessem, D. (1993). The complete T. rex. Simon & Schuster, New York City. Auch wenn diese Arme im Verhältnis zum Körper sehr klein sind und mit nur zwei Fingern recht nutzlos erscheinen, fielen bereits Osborn bei der Erstbeschreibung sehr große Muskelansatzflächen auf, was auf enorme Kraft hindeutet.53Osborn, H. F. & Brown, B. (1906). Tyrannosaurus, Upper Cretaceous carnivorous dinosaur. Bulletin of the AMNH, 22(16), 281–296. Die Armknochen von T. rex besitzen zudem außergewöhnlich dicke Knochenwände, was darauf hinweist, dass sie hohen Belastungen standhalten konnten. Der Bizeps eines ausgewachsenen T. rex könnte allein etwa 199 Kilogramm anheben. Gemeinsam mit anderen Muskeln hätte dies die Kraft der Armbeugung zusätzlich erhöht. Der Bizeps war dabei etwa 3,5-mal stärker als der eines gut trainierten Menschen. Schulter- und Ellenbogengelenk erlaubten allerdings nur etwa 40 beziehungsweise 45 Grad Bewegungsradius. Zum Vergleich: Bei Deinonychus lagen diese Werte bei etwa 88 und 130 Grad, beim Menschen kann sich der Arm an der Schulter nahezu vollständig drehen. Die Kombination aus massiven Knochen, starker Muskulatur und geringer Beweglichkeit könnte darauf hindeuten, dass die Arme eher dafür ausgelegt waren, Beute trotz heftiger Gegenwehr festzuhalten.54Carpenter, K. & Smith, M. (2001). Forelimb Osteology and Biomechanics of Tyrannosaurus rex. In Tanke, D. H. & Carpenter, K. (Hgg.), Mesozoic vertebrate life. Indiana University Press, Bloomington, 90–116. Steven M. Stanley schlug sogar vor, dass die Arme sogar als Waffen dienten und die Klauen dazu genutzt wurden, tiefe Wunden in die Beute zu reißen.55Pickrell, J. (2017). T. Rex’s Tiny Arms May Have Been Vicious Weapons. National Geographic.

Doch warum waren sie trotzdem so winzig? Eine interessante Hypothese wurde 2022 vorgestellt, die besagt, dass die Verkleinerung der Arme bei Tyrannosauriden wahrscheinlich keine spezielle Funktion erfüllte, sondern eine sekundäre Anpassung war, die auch bei vielen anderen Großtheropoden auftrat, wie bei Carcharodontosauriden und vor allem Abelisauriden, die sogar noch winzigere Ärmchen hatten als die Tyrannosauriden. Während Schädel und Kiefer immer größer und kräftiger wurden, seien die Arme kleiner geworden, um Verletzungen durch (versehentliche oder auch absichtliche!) Bisse anderer Tyrannosaurier zu vermeiden, wenn sie gemeinsam an einem Kadaver fraßen.56Padian, K. (2022). Why tyrannosaurid forelimbs were so short. An integrative hypothesis. Acta Palaeontologica Polonica, 67(1), 63–76. In meiner Romanreihe tragen die Arme des Männchens zudem einen auffälligen Federbesatz, der bei der Balz eine Rolle spielt. Diese Interpretation ist allerdings spekulativ.
Gleich- oder wechselwarm?
|
|
Bis in die zweite Hälfte des 20. Jahrhunderts hinein wurden Dinosaurier noch als wechselwarme, „kaltblütige“ Reptilien betrachtet. Doch ab den 1960er Jahren kamen daran während der sogenannten „Dinosaurier-Renaissanc“ Zweifel auf, vor allem aufgrund der Arbeiten von John Ostrom57Ostrom, J. H. (1969). Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Bulletin of the Peabody Museum of Natural History, 30. und Robert T. Bakker58Bakker, R. T. (1972). Anatomical and ecological evidence of endothermy in dinosaurs. Nature, 238(5359), 81–85. doi:10.1038/238081a059Bakker, R. T. (1986). The Dinosaur Heresies. Kensington Publishing, New York, 241.. Dabei wurde auch für T. rex ein endothermer („warmblütiger“) Stoffwechsel vorgeschlagen, was auf eine sehr aktive Lebensweise hindeuten würde. Histologische Untersuchungen der Knochen zeigen bei jungen T. rex sehr hohe Wachstumsraten, vergleichbar mit denen heutiger Säugetiere und Vögel. Wachstumskurven deuten außerdem darauf hin, dass das Wachstum ähnlich wie bei Vögeln und Säugetieren hauptsächlich auf die Jugendphase beschränkt war, statt lebenslang weiterzugehen wie bei vielen anderen wechselwarmen Wirbeltieren.60Horner, J. R. & Padian, K. (2004). Age and growth dynamics of Tyrannosaurus rex. Proceedings: Biological Sciences, 271(1551), 1875–1880. doi:10.1098/rspb.2004.2829 |


Auch Sauerstoffisotope in fossilisierten Knochen wurden genutzt, um Rückschlüsse auf die Körpertemperatur zu ziehen. Das Verhältnis bestimmter Sauerstoffisotope hängt mit der Temperatur zusammen, bei der der Knochen entstand. Bei einem T. rex-Exemplar zeigte sich zwischen den Wirbeln des Rumpfes und dem Schienbein nur ein Temperaturunterschied von etwa 4 bis 5 °C. Der Paläontologe Reese Barrick und der Geochemiker William Showers interpretierten dies als Hinweis darauf, dass T. rex eine relativ konstante innere Körpertemperatur besaß (Homöothermie) und sein Stoffwechsel zwischen dem moderner Reptilien und Säugetiere lag.61Barrick, R. E. & Showers, W. J. (1994). Thermophysiology of Tyrannosaurus rex. Evidence from Oxygen Isotopes. Science, 265(5169), 222–224. doi:10.1126/science.265.5169.222 Einen weiteren Ansatz nutzt die spektroskopische Untersuchung von Lipoxidations-Signalen, die als Nebenprodukte Stoffwechsels auftreten. Auch dies lieferte Hinweise darauf, dass T. rex seine Körpertemperatur konstant halten konnte, und dass ein endothermer Stoffwechsel sogar bei allen Dinosauriern ursprünglich angelegt war.62Wiemann, J. et al. (2022). Fossil biomolecules reveal an avian metabolism in the ancestral dinosaur. Nature, 606(7914), 522–526. doi:10.1038/s41586-022-04770-6 Einen weiteren Hinweis stellen die dorsotemporalen Fenster von T. rex dar, spezialisierte Öffnungen im Schädeldach, die ähnlich wie bei heutigen Krokodilen eine Rolle bei der Wärmeregulierung gespielt haben könnten.63Holliday, C. M. et al. (2019). The Frontoparietal Fossa and Dorsotemporal Fenestra of Archosaurs and Their Significance for Interpretations of Vascular and Muscular Anatomy in Dinosaurs. The Anatomical Record, 303(4), 1060–1074. doi:10.1002/ar.24218 Etwas Vorsicht ist bei dieser Interpretation aber trotzdem geboten. Denn selbst wenn T. rex tatsächlich eine konstante Körpertemperatur hatte, bedeutet dies jedoch nicht zwangsläufig, dass er sie vollständig selbst erzeugen konnte. Auch die sogenannte Gigantothermie erklärt dieses Phänomen: sehr große Tiere haben oft allein aufgrund ihrer Körpermasse die Fähigkeit, Wärme lange zu speichern, wie z.B. einige Meeresschildkröten.64Paladino, F. V., Spotila, J. R. & Dodson, P. (1999). A blueprint for giants. Modeling the physiology of large dinosaurs. In Farlow, J. O. & Brett-Surman, M. K. (Hgg.), The Complete Dinosaur. Indiana University Press, Bloomington, 491–504.65Chinsamy, A. & Hillenius, W. J. (2004). Physiology of nonavian dinosaurs. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The dinosauria. University of California Press, Berkeley, 643–659.66Seymour, R. S. (2013). Maximal Aerobic and Anaerobic Power Generation in Large Crocodiles versus Mammals. Implications for Dinosaur Gigantothermy. PLOS ONE, 8(7), e69361. doi:10.1371/journal.pone.0069361
Haut oder Federn?
Wenn T. rex einen gleichbleibenden Stoffwechsel besaß, stellt sich auch die Frage, ob seine Haut einen Isolationsschutz besessen haben könnte, um die Körperwärme auch langfristig zu speichern, oder auch Strukturen, um sie bei Überhitzung wieder abgeben zu können. Seit der Entdeckung von Federstrukturen bei fleischfressenden Dinosauriern wird auch bei T. rex immer wieder darüber spekuliert, ob er gefiedert gewesen sein könnte. Bei mehreren früheren Tyrannosauroiden aus China konnten Federn sogar bereits direkt nachgewiesen, z.B. bei Dilong paradoxus67Xing, X. et al. (2004). Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature, 431(7009), 680–684. doi:10.1038/nature02855 oder auch beim noch weit größeren Yutyrannus huali.68Xing, X. et al. (2012). A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature, 484(7392), 92–95. doi:10.1038/nature10906 Die bekannten Hautabdrücke von T. rex zeigen aber allesamt ein mosaikartiges Muster aus kleinen Schuppen, aber keine Ansatzstellen für Federkiele.69Bell, P. R. et al. (2017). Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution. Biology Letters, 13(6), 20170092. doi:10.1098/rsbl.2017.0092 Falls T. rex also Federn besaß, dann wohl nur in sehr begrenztem Ausmaß. So trug er wahrscheinlich kein vollständiges Federkleid: dies hätte einem so großen Tier in einem subtropischen Lebensraum einen Nachteil bei der Thermoregulierung verschafft.

Aus diesem Grund habe ich nur die Jungtiere von T. rex in Die Weißen Steine als gefiedert dargestellt. Mit zunehmenden Alter wird das Federkleid dann immer dünner. Die Erwachsenen haben es fast völlig verloren. Ledigich das Männchen nutzt noch eine „Mähne“ und Armfedern zur innerartlichen Kommunikation und bei der Balz.
Lippen?
Auch die Frage, ob die Zähne bei T. rex sichtbar blieben, wenn er sein rieisges Maul schloss, oder ob sie durch Lippen verborgen blieben, wird in der Forschung noch diskutiert. Ein Konferenzbeitrag aus dem Jahr 2016 schlug vor, dass Theropoden wie T. rex ihre oberen Zähne von Lippen bedeckt hatten, statt frei liegender Zähne wie bei heutigen Krokodilen. Grundlage dieser Hypothese war das Vorhandensein von Zahnschmelz, der laut Studie feucht gehalten werden muss. Dieses Problem ist bei wasserlebenden Tieren wie Krokodilen weniger relevant, weshalb ihre Zähne auch bei geschlossenem Maul sichtbar sein können.70Reisz, R. R. & Larson, D. (2016). Dental anatomy and skull length to tooth size ratios support the hypothesis that theropod dinosaurs had lips. 4th Annual Meeting, 2016, Canadian Society of Vertebrate Palaeontology. An diesem Vorschlag gibt es jedoch auch Kritik, und auch alternative hypothetische Ansätze. Eine Studie von 2017 schlug stattdessen vor, dass Tyrannosauriden große, flache Schuppen auf der Schnauze besaßen und keine Lippen, ähnlich wie moderne Krokodile.71Carr, T. D. et al. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7, 44942. doi:10.1038/srep44942 Allerdings besitzen Krokodile tatsächlich stark rissige, verhornte Haut und keine glatten Schuppen. Durch den Vergleich der rauen Knochenoberflächen von Tyrannosauriden mit der Hautstruktur heutiger Echsen kamen Forscher zu dem Schluss, dass Tyrannosauriden eher schuppenartige Haut ähnlich moderner Echsen hatten als eine krokodilartige Hautstruktur.72Milinkovitch, M. et al. (2013). Crocodile head scales are not developmental units but emerge from physical cracking. Science, 339(6115), 78–81. doi:10.1126/science.1226265
Die jüngsten Studien werfen aber immer mehr in die Waagschale für Lippen im Gesicht des T. rex, weshalb ich mich auch dazu entschlossen habe, ihn in meiner Romanreihe ebenfalls mit Lippen auszustatten. Im Jahr 2023 untersuchten Cullen und Kollegen anatomische Merkmale wie die Verteilung der kleinen Öffnungen (Foramina) in Gesicht und Kiefer, die stärker den Mustern moderner Schuppenkriechtiere wie etwa Waranen oder Meerechsen entsprachen. Diese Tiere haben Lippen. Ein Vergleich der Zähne von Daspletosaurus mit denen des Amerikanischen Alligators zeigte außerdem deutliche Unterschiede. Der Zahnschmelz der Tyrannosauriden zeigte kaum Abnutzung, während die Zähne moderner Krokodile auf der Außenseite stark abgerieben sind. Dies deutet darauf hin, dass die Zähne der Theropoden wahrscheinlich durch Lippen feucht gehalten wurden.73Cullen, T. M. et al. (2023). Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology. Science, 379(6639), 1348–1352. doi:10.1126/science.abo7877
Weichgewebe
 Mary H. Schweitzer Mary H. Schweitzer |
Ein gleichsam kurioser wie auch sehr aufregender Fakt zu T. rex ist: wir haben von ihm tatsächlich Weichgewebe überliefert, das noch nicht versteinert ist! In der Markhöhle eines fossilen Beinknochens des Exemplars „B-Rex“ (MOR 1125), der für den Abtransport absichtlich, wenn auch widerwillig zerbrochen werden musste, konnten Mary H. Schweitzer und ihr Team zu ihrer eigenen Überraschung zunächst einen ungewöhnlich fauligen Geruch und später tatsächlich weiches Gewebe feststellen. |
Die Forscher identifizierten flexible, verzweigte Blutgefäßstrukturen sowie faseriges, elastisches Gewebe, das der Knochenmatrix ähnelte. Zusätzlich fanden sie Mikrostrukturen, die roten Blutkörperchen ähnelten. Diese Strukturen erinnerten stark an Blutgefäße und Blutzellen heutiger Strauße. Auch wenn dies zuvor unmöglich erschien, können solche Strukturen bei ungewöhnlich guten Fossilisationsbedingungen auch nach Millionen von Jahren erhalten bleiben.74Schweitzer, M. H. et al. (2005). Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science, 307(5717), 1952–1955. doi:10.1126/science.1108397 Inzwischen konnte auch an zwei weiteren Tyrannosauriern und einem Hadrosaurier Weichgewebe nachgewiesen werden. Untersuchungen deuten darauf hin, dass Tyrannosaurier eng mit Vögeln verwandt sind. Auch die ursprüngliche chemische Zusammensetzung des Knochens blieb teilweise erhalten, darunter Elemente wie Schwefel, Kalzium und Zink, die mit Knochenumbauprozessen zusammenhängen und Ähnlichkeiten zu modernen Vogelknochen zeigen.75Anné, J. et al. (2023). Independent Evidence for the Preservation of Endogenous Bone Biochemistry in a Specimen of Tyrannosaurus rex. Biology, 12(2), 264. doi:10.3390/biology12020264
Die angeblichen Weichgewebereste wurden jedoch auch angezweifelt. Einige Forscher argumentierten, dass es sich in Wirklichkeit um schleimige Biofilme von modernen Bakterien handle, die die Hohlräume ehemaliger Blutgefäße und Zellen ausgekleidet hätten. Die vermeintlichen Blutkörperchen seien tatsächlich sogenannte Framboide, also mikroskopisch kleine eisenhaltige Mineralkügelchen. Solche Strukturen fanden die Forscher auch in anderen Fossilien, darunter sogar in einem Ammoniten, wo sie offensichtlich nichts mit Blut zu tun haben konnten.76Kaye, T. G. et al. (2008). Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms. PLOS ONE, 3(7), e2808. doi:10.1371/journal.pone.0002808 Schweitzer kritisierte diese Interpretation scharf. Sie argumentierte, dass bislang kein Biofilm bekannt sei, der verzweigte hohle Röhren wie jene aus ihrer Studie erzeugen könne. 2011 veröffentlichte sie mit ihrem Team eine weitere Analyse der geborgenen Kollagenreste. Dabei zeigte sich, dass vor allem die inneren Bereiche der Kollagenfasern erhalten geblieben waren, genau jene Teile, die theoretisch am ehesten eine lange Proteinzerstörung überstehen könnten.77San Antonio, J. D. et al. (2011). Dinosaur Peptides Suggest Mechanisms of Protein Survival. PLOS ONE, 6(6), e20381. doi:10.1371/journal.pone.0020381 Andere Studien widersprachen ebenfalls der Biofilm-Hypothese und bestätigten die Existenz verzweigter gefäßähnlicher Strukturen innerhalb fossiler Knochen.78Peterson, J. E., Lenczewski, M. E. & Scherer, R. P. (2010). Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs. PLOS ONE, 5(10), e13334. doi:10.1371/journal.pone.0013334
Geschlechtsdimorphismus?
Als die Zahl der bekannten T. rex-Fossilien zunahm, begannen Wissenschaftler die Unterschiede zwischen einzelnen Individuen genauer zu untersuchen. Dabei schienen sich zwei unterschiedliche Körpertypen, sogenannte Morphe, abzuzeichnen, ähnlich wie bei einigen anderen Theropodenarten. Einer dieser Morphen war kräftiger und robuster gebaut, der andere schlanker und graziler. Dies wurde Anfang der 1990er als Geschlechtsdimorphismus gedeutet, also dass einer der Morphen das Männchen, der andere das Weibchen gewesen sei. Der robuste Morph wurde dabei als weiblich angesehen. Das vermeintlich breitere Becken hätte demnach die Eiablage erleichtert.79Carpenter, K. (1992). Variation in Tyrannosaurus rex. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Approaches and Perspectives. Cambridge University Press, Cambridge, 141–145. Außerdem kam die These auf, dass robuste Tiere einen verkleinerten Chevron-Knochen am ersten Schwanzwirbel besaßen, was angeblich ebenfalls mit der Eiablage zusammenhing, was man damals auch bei Krokodilen annahm.80Larson, P. L. (1994). Tyrannosaurus sex. In Rosenberg, G. D. & Wolberg, D. L. (Hgg.), Dino Fest. The Paleontological Society Special Publications, 139–155.
In den letzten Jahren wurden die Hinweise auf Geschlechtsdimorphismus jedoch immer mehr in Zweifel gezogen. Eine Studie von 2005 zeigte, dass frühere Behauptungen über geschlechtsspezifische Unterschiede der Chevron-Knochen bei Krokodilen falsch waren.81Erickson, G. M., Kristopher, L. A. & Larson, P. (2005). Androgynous rex. The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs. Zoology, 108(4), 277–286. doi:10.1016/j.zool.2005.08.001 Zusätzlich wurde bei „Sue“, dem damals robustesten T. rex-Exemplar überhaupt, ein vollständig entwickelter Chevron-Knochen am ersten Schwanzwirbel entdeckt. Dieses Merkmal eignete sich somit nicht zur Geschlechtsbestimmung. Da T. rex-Fossilien von Saskatchewan bis New Mexico gefunden wurden, könnten die Unterschiede zwischen einzelnen Individuen stattdessen auch geografische Variationen widerspiegeln. Ebenso ist möglich, dass robuste Tiere einfach ältere oder schlicht besonders kräftige Individuen waren.82Brochu, C. R. (2003). Osteology of Tyrannosaurus rex. Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Society of Vertebrate Paleontology Memoirs, 7, 1–138. doi:10.2307/3889334
Bis heute konnte nur ein einziges T. rex-Exemplar eindeutig einem Geschlecht zugeordnet werden. Untersuchungen am Exemplar „B-rex“ (MOR 1125) zeigten den Erhalt von Weichgewebe in mehreren Knochen. Ein Teil dieses Gewebes wurde als medulläres Knochengewebe identifiziert. Dieses Gewebe bildet sich bei heutigen Vögeln nur dann, wenn ein Tier kurz vor der Eiablage steht, da es als Kalziumspeicher für die Eierschalen dient. Dies spricht stark dafür, dass „B-rex“ ein weibliches Tier gewesen sein muss.83Schweitzer, M. H., Wittmeyer, J. L. & Horner, J. R. (2005). Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. Science, 308(5727), 1456–1460. doi:10.1126/science.1112158 Bei Krokodilen kommt diese Gewebestruktur dagegen nicht vor.84Schweitzer, M. H. et al. (2007). Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? Bone, 40(4), 1152–1158. doi:10.1016/j.bone.2006.10.029
In meiner Romanreihe habe ich mich dafür entschieden, trotzdem einen Geschlechtsdimorphismus für T. rex zu zeigen, und habe das Weibchen größer gemacht als das Männchen. Derartige Geschlechtsunterschiede sind auch bei heutigen Raubvögeln häufig. Außerdem konnte ich so eine gewisse Dramatik bei den Familienstrukturen von T. rex erzählerisch aufbauen. Die wissenschaftlichen Hinweise dazu sind zwar nach wie vor gegeben und plausibel, auch wenn harte Evidenzen dazu leider fehlen.

Oder sogar mehrere Arten?
 Gregory S. Paul Gregory S. Paul |
In einer Studie aus dem Jahr 2022 griffen Gregory S. Paul und Kollegen die These der mehreren Morphen wieder auf, argumentierten jedoch, dass T. rex im traditionellen Sinne tatsächlich aus drei verschiedenen Arten bestehe. Die Aufteilung in mehrere Arten begründeten sie vor allem mit der starken Variation in den Proportionen und der Robustheit der Oberschenkelknochen sowie anderer Skelettelemente verschiedener T. rex-Exemplare. Zusätzlich wurden die angenommenen robusten und grazilen Morphotypen sowie die Anzahl kleiner schneidezahnartiger Zähne im Dentarium als Hinweise herangezogen.85Paul, G. S., Persons IV, W. S. & van Raalte, J. (2022). The Tyrant Lizard King, Queen and Emperor. Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus. Evolutionary Biology, 49(2), 156–179. doi:10.1007/s11692-022-09561-5 |
Paul und Kollegen definierten die Arten folgendermaßen:
- T. rex besitze eine robuste Anatomie, ein mittleres Verhältnis von Femurlänge zu Umfang sowie einen einzelnen schlanken schneidezahnartigen Zahn im Unterkiefer. Das Referenzexemplar ist das Typusexemplar im Carnegie Museum, CM 9380.
- T. imperator sei besonders robust, mit einem niedrigen Verhältnis von Femurlänge zu Umfang und zwei dieser schlanken Zähne. Er wäre stratigraphisch die älteste Art, aus der sich die anderen beiden im Laufe der Zeit entwickelt haben könnten. Das Referenzexemplar dieser Spezies sei „Sue“ (FMNH PR 2081).
- T. regina dagegen sei graziler gebaut, mit einem hohen Femurverhältnis und nur einem schneidezahnartigen Zahn. Das referenzexemplar ist „Wankel Rex“ (MOR 555).
 Thomas D. Carr Thomas D. Carr |
Mehrere führende Paläontologen, vor allem Thomas D. Carr, kritisierten die Studie jedoch oder äußerten starke Zweifel an ihren Schlussfolgerungen. Ihre Kritik wurde später auch in einem wissenschaftlichen Gegen-Paper veröffentlicht. Sie halten es zwar grundsätzlich für möglich, dass mehr als eine Tyrannosaurus-Art existierte, betrachteten die vorgelegten Daten jedoch als unzureichend, um die vorgeschlagenen Arten zu stützen.86Carr, T. D. et al. (2022). Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America. A Comment on „The Tyrant Lizard King, Queen and Emperor. Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus“. Evolutionary Biology, 49(3), 314–341. doi:10.1007/s11692-022-09573-1 |
Gregory S. Paul hält jedoch weiterhin an seiner These fest und veröffentlichte 2025 ein Gegen-Gegen-Paper. Er argumentierte, dass sich diese vorgeschlagenen Arten zusätzlich anhand der Form der Postorbitalhöcker, also knöcherner Verdickungen hinter den Augen, unterscheiden ließen.87Paul, G. S. (2025). A presentation of the current data on the exceptionally diverse non-tyrannosaurid eutyrannosaur and Tyrannosaurini genera and species of western North America during the End Cretaceous North American Interchange. Mesozoic, 2(2), 085–138. doi:10.11646/mesozoic.2.2.1 Paul steht mit seinen Überlegungen allerdings inzwischen weitestgehend isoliert da.
Tyrannosaurus mcreaensis und die mögliche Herkunft von T. rex
 |
Noch eine weitere Tyrannosaurus-Art wird in der Forschung diskutiert, und das bereits seit über vier Jahrzehnten. Bereits 1983 entdeckten Forscher in der McRae-Formation in New Mexico Teile des Schädels, des Unterkiefers, isolierte Zähne und Chevron-Knochen eines großen Theropoden. Das Material wurde zunächst Tyrannosaurus rex zugeschrieben.88Lozinsky, R. P., Hunt, A. P., Wolberg, D. L. & Lucas, S. G. (1984). Late Cretaceous (Lancian) dinosaurs from the McRae Formation, Sierra County, New Mexico. New Mexico Geology, 6(4), 72–77. doi:10.58799/NMG-v6n4.72 |
1986 wurde es näher beschrieben, aber ebenfalls noch als sehr urtümlicher T. rex.89Gillette, D. D., Wolberg, D. L. & Hunt, A. P. (1986). Tyrannosaurus rex from the McRae Formation (Lancian, Upper Cretaceous), Elephant Butte reservoir, Sierra County, New Mexico. New Mexico Geological Society Guidebook, 37, 235–238. doi:10.56577/FFC-37.235 Lehman und Carpenter schlugen 1990 vor, dass das Fossil zu einer neuen Tyrannosauriden-Gattung gehören könnte90Lehman, T. M. & Carpenter, K. (1990). A partial skeleton of the tyrannosaurid dinosaur Aublysodon from the Upper Cretaceous of New Mexico. Journal of Paleontology, 64(6), 1026–1032. doi:10.1017/S0022336000019843, während Carr und Williamson dies 2000 aber zurückwiesen.91Carr, T. D. & Williamson, T. E. (2000). A review of Tyrannosauridae (Dinosauria: Coelurosauria) from New Mexico. Bulletin, 17. New Mexico Museum of Natural History and Science, 113–145. Sullivan und Lucas argumentierten 2015, dass es nur wenige Belege dafür gebe, dass es sich tatsächlich um Tyrannosaurus rex handle, und klassifizierten das Fossil deshalb vorsichtig als cf. Tyrannosaurus sp. Sie vermuteten außerdem, dass der sogenannte „McRae-Tyrannosaurier“ deutlich vor der Zeit von T. rex lebte, also älter als 68 Millionen Jahre sei.92Sullivan, R. M. & Lucas, S. G. (2015). Cretaceous Vertebrates of New Mexico. New Mexico Museum of Natural History and Science Bulletin, 68.
Dalman und Kollegen untersuchten das Exemplar 2024 erneut und kamen zu dem Schluss, dass es sich tatsächlich um eine eigene, deutlich ältere Art von Tyrannosaurus handeln müsse, womit sie der These neues Gewicht verliehen, dass T. rex sich ursprünglich auf dem nordamerikanischen Kontinent entwickelte, und nicht, wie andere vermuteten, als invasive Spezies aus Asien eingewandert sei. Das Exemplar schätzten sie auf etwa 12 Meter Länge, womit es ähnlich groß wie ein ausgewachsener T. rex wäre. Unterschiede zwischen beiden Formen betreffen vor allem den Schädel: Das Dentarium von T. mcraeensis ist verhältnismäßig länger, besitzt ein weniger ausgeprägtes „Kinn“ und der Unterkiefer ist flacher, was auf eine schwächere Beißkraft hindeuten könnte. Auch die Zähne sind stumpfer und seitlich stärker zusammengedrückt, während die Postorbitalhöcker weniger stark ausgeprägt sind. Zudem weist die Skelettanatomie Gemeinsamkeiten mit Tarbosaurus und Zhuchengtyrannus auf. Aufgrund dieser Unterschiede, aber auch des großen angenommen zeitlichen Abstand zu T. rex, gaben die Forscher dem Exemplar den neuen Namen Tyrannosaurus mcraeensis, benannt nach der Formation, in wecher er gefunden worden war. Diese ist einige Millionen Jahre älter als die klassische Zeitspanne von T. rex, der am Ende des Maastrichtiums lebte, und wird auf ein Alter zwischen 72,7 und 70,9 Millionen Jahren datiert, also noch aufs spätes Campanium bis frühestes Maastrichtium. 93Dalman, S. G. et al. (2024). A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism. Scientific Reports, 13(1), 22124. doi:10.1038/s41598-023-47011-0

Ein Konferenzabstract aus demselben Jahr kam jedoch zu einem jüngeren Alter: Die genaue Fundschicht von T. mcraeensis sei zwischen 69,0 und 66 Ma alt. Damit läge das Alter deutlich näher an T. rex als ursprünglich angenommen.94Schantz, E. & Amato, J. M. (2024). Dating dinosaurs in the Laramide foreland. U-Pb geochronologic constraints on a stratigraphic section containing Alamosaurus and the postulated Tyrannosaurus mcraeensis in the Love Ranch Basin. New Mexico Geological Society of America Abstracts with Programs, 56, 9.95Scherer, C. R. (2025). Multiple lines of evidence support anagenesis in Daspletosaurus and cladogenesis in derived tyrannosaurines. Cretaceous Research, 169, 106080. doi:10.1016/j.cretres.2025.106080 Nicht zuletzt deshalb wurde die Gültigkeit von T. mcraeensis bereits 2025 von anderen Forschern infrage gestellt. Sie argumentierten, dass die Verwendung von Alamosaurus-Fossilien, die als biostratigraphische Marker zur Altersbestimmung herangezogen wurden, problematisch sei, da die Sedimentationsraten stark schwanken könnten. Außerdem bezweifelten sie, dass sich die Merkmale des Holotypus zuverlässig außerhalb der bekannten Variationsbreite von T. rex bewegen. Unabhängig von Alter und Gültigkeit von T. mcraeensis kamen die Autoren zu dem Schluss, dass die Gattung Tyrannosaurus zwar in Laramidia entstand, ihre Vorfahren aber wohl doch aus Asien eingewandert waren.96Morrison, C. et al. (2025). Rise of the king. Gondwanan origins and evolution of megaraptoran dinosaurs. Royal Society Open Science, 12(5), 242238. doi:10.1098/rsos.242238 Später im selben Jahr betrachteten auch Voris und Kollegen T. mcraeensis als jüngeres Synonym von T. rex. Sie stimmten den Bedenken bezüglich der stratigraphischen Daten zu und kamen zu dem Ergebnis, dass alle vorgeschlagenen diagnostischen Merkmale innerhalb der bekannten Variationsbreite von T. rex liegen.97Voris, J. T. et al. (2025). A new Mongolian tyrannosauroid and the evolution of Eutyrannosauria. Nature, 642(8069), 973–979. doi:10.1038/s41586-025-08964-6 Auch Thomas D. Carr erkannte T. mcraeensis in seiner Untersuchung des Holotyp-Schädels von T. rex nicht als gültige Art an und interpretierte die vorgeschlagenen Unterschiede ebenfalls als normale Variation innerhalb von T. rex.98Carr, T. D. (2025). Observations on the skull of the type specimen of Tyrannosaurus rex Osborn, 1905. All Earth, 37(1), 2539638, 1–66. doi:10.1080/27669645.2025.2539638 An der Gültigkeit von „T-Mac“ bestehen inzwischen ebenso Zweifel wie an T. regina und T. imperator. Und der Streit um die Herkunft von T. rex, also die Frage, ob seine Vorfahren aus Nordamerika oder doch aus Asien kamen, ist somit ebenfalls noch offen.
Sinnesleistungen
Eine der wohl häufigsten Falschannahmen über T. rex hat sich wahrscheinlich durch Michael Crichtons Buch Jurassic Park (1989) bzw. die Verfilmung durch Steven Spielberg (1993) in den Köpfen vieler Menschen festgesetzt. Dort ist der Tyrannosaurus nur imstande, sich bewegende Objekte zu erkennen, da seine visuelle Wahrnehmung offenbar wie die eines Frosches kalibriert sei. Die Figuren Alan Grant und Lex Murphy können die Begegnung mit einem T. rex nur knapp überleben, indem sie reglos stehenbleiben.
Doch könnte diese Darstellung von der Wahrheit wohl kaum weiter entfernt sein! Mehrere Fachpublikationen kamen zu dem Ergebnis, dass Tyrannosaurus über ein außergewöhnlich gutes Sehvermögen verfügte. Mithilfe rekonstruierter Schädelmodelle verschiedener Dinosaurier berechnete Stevens für Tyrannosaurus ein binokulares Sichtfeld von etwa 55°, was mehr ist als bei heutigen Greifvögeln. Außerdem schätzte er die Sehschärfe auf etwa das 13-fache eines Menschen, was ebenfalls deutlich höher als die eines Adlers ist, der immerhin das 3,6-fach so scharf sehen kann wie ein Mensch. Stevens ging davon aus, dass Tyrannosaurus (auch unbewegte!) Objekte noch aus bis zu 6 Kilometern Entfernung scharf erkennen konnte.99Stevens, K. A. (2006). Binocular vision in theropod dinosaurs. Journal of Vertebrate Paleontology, 26(2), 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2100Jaffe, E. (2006). Sight for Saur Eyes. T. rex vision was among nature’s best. Science News, 170(1), 3–4. doi:10.2307/4017288 Thomas Holtz Jr. vermutete, dass dieses ausgeprägte räumliche Sehvermögen mit der Beute von Tyrannosaurus zusammenhing. Tiere wie Triceratops, Ankylosaurus oder große Hadrosaurier waren gefährliche Gegner, weshalb präzise Angriffe entscheidend gewesen sein könnten. Holtz formulierte dies sinngemäß so:
„Die Jagd eines Tyrannosaurus können wir uns so vorstellen: herankommen, den entscheidenden Treffer landen und das Tier zu Boden bringen.“
Konträr dazu hatte z.B. Acrocanthosaurus eben keine so ausgeprägte räumliche Wahrnehmung, da er sich wohl eher auf Sauropoden als Beute spezialisiert hatte.101Holtz, T. R. Jr. (2013). The Life and Times of Tyrannosaurus rex, with Dr. Thomas Holtz. Lecture, Burke Museum of Natural History and Culture, Seattle, WA. Wie gut ein Dinosaurier bei Nacht oder im Dämmerlicht sehen konnte, lässt sich außerdem gut anhand sogenannter Skleralringe untersuchen. Je größer der Ring, umso mehr Licht kann er hereinlassen, weshalb sehr große Skleralringe eher bei nachtaktiven Tieren vorkommen. Obwohl bislang kein solcher Skleralring von Tyrannosaurus gefunden wurde, rekonstruierte Kenneth Carpenter dessen Größe anhand von Gorgosaurus. Seine Berechnungen deuten darauf hin, dass Tyrannosaurus wohl eher nicht so gut bei schwachem Licht sehen konnte und wahrscheinlich hauptsächlich tagsüber jagte.102Carpenter, K. (2013). A Closer Look at the Scavenging versus Predation by Tyrannosaurus rex. In Parrish, M. J., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. Life of the Past. Indiana University Press, Bloomington, 265–278.
Eine Studie von Lawrence Witmer und Ryan Ridgely von der Ohio University zeigte, dass T. rex die stark ausgeprägten Sinnesleistungen anderer Coelurosaurier teilte. Dazu gehörte zum einen ein außerordentlich gutes Hörvermögen. Die Cochlea im Innenohr von T. rex war ungewöhnlich lang, was dafür spricht, dass Tyrannosaurus besonders gut tieffrequente Geräusche wahrnahm. Solche tiefen, weithallenden Laute spielten wahrscheinlich eine wichtige Rolle im Verhalten und bei der Kommunikation der Tiere. Außerdem erlaubten Anatomie und Neurologie schnelle und koordinierte Bewegungen von Augen und Kopf, wodurch Beutebewegungen über größere Entfernungen erkannt werden konnten, sowie ein ausgezeichneter Geruchssinn. 103Witmer, L. M. & Ridgely, R. C. (2009). New Insights into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior. The Anatomical Record, 292(9), 1266–1296. doi:10.1002/ar.20983 T. rex besaß im Verhältnis zur Gehirngröße extrem große Riechkolben und hochsensible Riechnerven, die zu den größten überhaupt bekannten bei Nichtvogel-Dinosauriern zählen.104Zelenitsky, D. K., Therrien, F. & Kobayashi, Y. (2008). Olfactory acuity in theropods. Palaeobiological and evolutionary implications. Proceedings of the Royal Society B: Biological Sciences, 276(1657), 667–673. doi:10.1098/rspb.2008.1075

2017 zeigten Thomas D. Carr und Kollegen anhand zahlreicher kleiner Öffnungen in den Gesichtsknochen verwandter Tyrannosauriden, dass die Schnauze wahrscheinlich sehr empfindlich war. Die Forscher vermuteten, dass Tyrannosaurier ihre Schnauze möglicherweise ähnlich wie heutige Krokodile nutzten, vielleicht um die Temperatur von Nestern zu prüfen oder Eier und Jungtiere vorsichtig zu bewegen.105Carr, T. D. et al. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7, 44942. doi:10.1038/srep44942 Eine weitere Studie von 2021 deutete ebenfalls auf einen ausgeprägten Tastsinn hin. Demnach könnte Tyrannosaurus mithilfe empfindlicher Nervenkanäle in den Kiefern sogar feinste Unterschiede in Material und Bewegung gespürt haben. Dies hätte dem Tier erlaubt, verschiedene Teile eines Kadavers unterschiedlich zu bearbeiten. Die empfindliche Schnauze könnte außerdem bei Nestbau, Brutpflege oder sozialer Kommunikation eine Rolle gespielt haben.106Kawabe, S. & Hattori, S. (2021). Complex neurovascular system in the dentary of Tyrannosaurus. Historical Biology, 34(7), 1137–1145. doi:10.1080/08912963.2021.1965137 Neuere Untersuchungen relativieren diese Interpretation allerdings teilweise. Sie weisen darauf hin, dass besonders dichte Netzwerke solcher Nervenkanäle häufiger bei wasserlebenden Tieren oder Arten mit Hornschnäbeln vorkommen. Die besagten Kanäle von Tyrannosaurus erscheinen zwar komplexer als bei vielen anderen Dinosauriern, könnten aber eher durchschnittlich empfindlich gewesen sein. Möglich ist daher auch, dass diese Strukturen vor allem der Wärmeregulierung oder optischen Signalen dienten.107Bouabdellah, F., Lessner, E. & Benoit, J. (2022). The rostral neurovascular system of Tyrannosaurus rex. Palaeontologia Electronica, 25(1), 1–20. doi:10.26879/1178
Auch die relative Gehirngröße von Tyrannosaurus wurde bereits untersucht und erste Einschätzungen zur möglichen Intelligenz vorgenommen. Die Ergebnisse zeigten, dass T. rex im Verhältnis zu seiner Körpergröße eines der größten Gehirne aller erwachsenen Nichtvogel-Dinosaurier besaß, im Vergleich zu seiner Gesamtmasse war es dagegen winzig und wog nur etwa 400 Gramm.108Brochu, C. A. (2000). A digitally-rendered endocast for Tyrannosaurus rex. Journal of Vertebrate Paleontology, 20(1), 1–6. doi:10.1671/0272-4634(2000)020[0001:ADREFT]2.0.CO;2 Allerdings lässt die Größe allein nicht unbedingt direkte Schlüsse auf die Intelligenz des Tieres zu. Bei Säugetieren ist z.B. vielmehr der Enzephalisationsquotient (EQ) ausschlaggebend, eine Maßeinheit, welche die relative Gehirngröße eines Tieres im Verhältnis zu seiner Körpergröße bewertet. T. rex besaß tatsächlich einen der höchsten bekannten EQ-Werte aller Nichtvogel-Dinosaurier, der bei etwa 47,5 und 49,53 Prozent lag. Nur bei einigen kleineren Maniraptoren war der EQ noch höher. Tatsächlich liegt T. rex damit etwa im Bereich einiger heutiger Vögel, am ehesten passen seine Werte jedoch zu denen eines Alligators, der auf 45,9 bis 47,9 Prozent kommt.109Hurlburt, G. S., Ridgely, R. C. & Witmer, L. M. (2013). Relative size of brain and cerebrum in Tyrannosaurid dinosaurs. An analysis using brain-endocast quantitative relationships in extant alligators. In Parrish, M. J., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. Indiana University Press, 134–154.

 Stephen L. Brusatte Stephen L. Brusatte |
Andere Forscher haben die Intelligenz von T. rex allerdings auch bereits deutlich höher eingeschätzt. Dem vielbeachteten Werk Aufstieg und Fall der Dinosaurier (2018) von Stephen L. Brusatte zufolge lag sein EQ in einem vergleichbaren Bereich wie der eines Schimpansen, wobei darin allerdings durchaus angemerkt wurde, dass die EQs von Säugetieren und Reptilien nur begrenzt miteinander vergleichbar seien.110Brusatte, S. (2018). The Rise and Fall of the Dinosaurs. HarperCollins Publishers, New York, NY, 219. |
Der neuronale Aufbau des Gehirns unterscheidet sich bei beiden Tiergruppen mitunter sehr stark. Aufschluss über eine mögliche Intelligenz gibt somit nicht allein der EQ, auch die schiere Anzahl der Neuronen (Nervenzellen) ist ein wichtiger Faktor, sowie ihr grad an synaptischer Verknüpfung. Auch dazu gab es bereits Schätzungen.
 Suzana Herculano-Houzel Suzana Herculano-Houzel |
Eine medial viel beachtete (und teils auch sehr bespöttelte!) Studie veröffentlichte Suzana Herculano-Houzel 2023, die zu dem Ergebnis kam, dass T. rex bis zu drei Milliarden Neuronen besaß und damit mindestens so intelligent wie ein Pavian gewesen sei. Demnach wäre es T. rex eventuell sogar möglich gewesen, einfachste Werkzeuge zu nutzen und sein Wissen an die nächsten Generationen weiterzugeben.111Herculano-Houzel, S. (2023). Theropod dinosaurs had primate-like numbers of telencephalic neurons. Journal of Comparative Neurology, 531(9), 962–974. doi:10.1002/cne.25453 |
Doch schon ein Jahr später meldeten sich die meisten führenden T. rex-Experten mit einem umfangreichen Gegenpaper zu Wort, die viele der neurologischen Resultate zurückweisen und klar widerlegen konnten. Zugleich lieferten sie nun eine überarbeitete Schätzung des EQs und der Neuronendichte, mit deutlich konservativeren Werten. Es sind aber, das sagte auch das Forscherteam, noch viele Studien und auch mehr aussagekräftigeres Material notwendig, um gültige Aussagen über die Intelligenz von T. rex zu treffen, wie auch über seine möglichen Fähigkeiten und auch seine Lebensweise.112Caspar, K. R. et al. (2024). How smart was T. rex? Testing claims of exceptional cognition in dinosaurs and the application of neuron count estimates in palaeontological research. The Anatomical Record, 307(12), 3685–3716. doi:10.1002/ar.25459
Sozialverhalten
Direkt an die Frage nach der möglichen Intelligenz von T. rex knüpft sich eine weitere an, nämlich die, was er damit anfangen konnte und wie er insbesondere damit mit seinen Artgenossen interagierte. Allerdings geben uns auf letztere Frage nun nicht unbedingt nur neurologische Untersuchungen eine Antwort, sondern eher die Zusammenhänge der Fossilien selbst sowie auch Spuren.
 Philip J. Currie Philip J. Currie |
Philip J. Currie schlug Ende der 1990er vor, dass Tyrannosaurus möglicherweise in Gruppen jagte. Dabei verglich er T. rex mit verwandten Arten wie Tarbosaurus bataar und Albertosaurus sarcophagus und verwies auf sogenannte Bone Beds, also Funde von mehreren Individuen an ein uns derselben Stelle, die auf ein geselliges Verhalten hindeuten.113Currie, P. J. (1998). Possible evidence of gregarious behaviour in tyrannosaurids. Gaia, 15, 271–277. |
Curries Hypothese einer Rudeljagd bei T. rex wurde jedoch kritisiert, da sie nie in einer begutachteten Fachpublikation veröffentlicht wurde, sondern hauptsächlich in einem Fernsehinterview und dem Buch Dino Gangs diskutiert wurde, sowie dass er als „Beweis“ nur mögliche Interpretationen bei anderen Tyrannosauriern vorbrachte und nicht bei T. rex selbst. Zusätzlich sind solche Gemeinschaftsfunde auch anders zu erklären, etwa durch Naturkatastrophen oder Prädatorenfallen. Currie räumte die Schwächen bei seinen Überlegungen in der Folge auch selbst ein. Andere Forscher vermuteten auch, dass einige dieser Funde eher ein Verhalten ähnlich dem heutiger Komodowarane widerspiegeln könnten. Diese versammeln sich auch ohne differenziertes Sozialverhalten oft trotzdem in großer Zahl, etwa wenn sie gemeinsam an einem Kadaver fressen. Manche Wissenschaftler gehen daher so weit zu sagen, dass echte Rudeljagd bei Dinosauriern möglicherweise überhaupt nicht existierte, da so ein komplexes organisiertes Verhalten selbst bei heutigen Raubtieren vergleichsweise selten ist.114Roach, B. T. & Brinkman, D. T. (2007). A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. Bulletin of the Peabody Museum of Natural History, 48(1), 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2
Ein Fund aus South Dakota, bei dem drei T. rex-Skelette dicht beieinander entdeckt wurden, könnte aber tatsächlich auf ein Rudel oder wenigstens einen losen Sozialverband hinweisen.115Collins, N. (2011). Tyrannosaurus rex hunted in packs. The Telegraph. Eines der Skelette, offenbar noch ein Jungtier, steht heute im Museum des Dinoparks im Altmühltal und trägt den Spitznamen „Rocky“. Weitere mögliche Hinweise auf ein Gruppenverhalten von Tyrannosauriden stammen von fossilen Fährten aus der Wapiti Formation im Nordosten von British Columbia in Kanada. Dort wurden die Spuren von drei Tyrannosauriden entdeckt, die offenbar alle gemeinsam in dieselbe Richtung liefen.116McCrea, R. T. (2014). A Terror of Tyrannosaurs. The First Trackways of Tyrannosaurids and Evidence of Gregariousness and Pathology in Tyrannosauridae. PLOS ONE, 9(7), e103613. doi:10.1371/journal.pone.0103613 Nicht zuletzt deutet auch die überwältigende Zahl an Nachweisen für Verletzungen an Fossilien von T. rex, die sehr wahrscheinlich durch Artgenossen entstanden, auf regelmäßige Auseinandersetzungen mit diesen hin. Das Exemplar „Wyrex“(HMNS 2006.1743.01 ) besitzt beispielsweise ein Loch im Jochbein sowie schwere Verletzungen am Schwanz, die Anzeichen einer Knochenheilung zeigen.117Rothschild, B. M. (2013). Clawing Their Way to the Top. Tyrannosaurid Pathology and Lifestyle. In Parrish, M. J., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. Life of the Past. Indiana University Press, Bloomington, 211–222.118Anné, J. et al. (2023). Unusual lesions seen in the caudals of the hadrosaur, Edmontosaurus annectens. The Anatomical Record, 306(3), 594–606. doi:10.1002/ar.25078
In jedem Fall könnte eine koordinierte Gemeinschaftsjagd sehr effizient gewesen sein, um wehrhafte Beutetiere wie Edmontosaurus, Triceratops oder Ankylosaurus zu überwältigen. Aus diesem Grund habe ich mich auch dazu entschlossen, in Die Weißen Steine ein komplexes Sozialverhalten bei T. rex zu zeigen. Allerdings ist dieses weit weniger „harmonisch“ als etwa in einem Wolfs- oder Löwenrudel, sondern eher als lockerer Sozialverband konzipiert, wie ihn viele heutige Vögel und Reptilien zeigen. Es gibt außerdem starke Hinweise darauf, dass T. rex wenigstens gelegentlich Kannibalismus betrieb. Bei einigen Exemplaren wurden Zahnspuren an Fußknochen, Oberarm und Mittelfuß eines anderen Individuums gefunden, also an Körperstellen, die eher unwahrscheinlich bei einem innerartlichen Kampf gebissen worden sein könnten. Die Struktur der Verletzungen deutet eher darauf hin, dass hier ein T. rex erst nach dem Tod seines Artgenossen an dessen Kadaver gefressen hat.119Longrich, N. R. et al. (2010). Cannibalism in Tyrannosaurus rex. PLOS ONE, 5(10), e13419. doi:10.1371/journal.pone.0013419 Die Kannibalismus-These habe ich auch in Band IV von Die Weißen Steine verarbeitet.

Geschwindigkeit
Noch ein weitere Mythos, den es zu entkräften gilt, rankt sich um die mögliche Höchstgeschwindigkeit, mit der T. rex seine Beute verfolgen konnte. Wohl jedem Kino- als auch Dino-Fan ist sicherlich die Szene aus Jurassic Park im Gedächtnis geblieben, in welcher der ausgebrochene T. rex einen Jeep jagt und ihn beinahe einholt. Doch war dies wirklich möglich?
Wissenschaftler haben im Laufe der letzten Jahre sehr unterschiedliche Schätzungen zur möglichen Höchstgeschwindigkeit von T. rex ermittelt. Forscher müssen sich bei solchen Berechnungen auf Modelle stützen, weil zwar zahlreiche Spuren großer Theropoden bekannt sind, aber keine eindeutig zeigt, dass diese Tiere tatsächlich rannten, als sie diese hinterließen. Wie die meisten heuitgen Tiere rannten Dinosaurier nur in ganz bestimmten Lebenssituationen in ihrer höchstmöglichen Geschwindigkeit, etwa wenn sie jagten oder flohen. Den größten Teil ihres Lebens bewegten sie sich aber in deutlich langsameren Gangarten.120Hutchinson, J. R. (2004). Biomechanical modeling and sensitivity analysis of bipedal running ability. II. Extinct taxa. Journal of Morphology, 262(1), 441–461. doi:10.1002/jmor.10240 Als extrem massiver und schwer gebauter Fleischfresser konnte er vermutlich nicht so schnell laufen wie andere große Theropoden wie Carnotaurus oder Giganotosaurus, die mit 45km/h tatsächlich sehr schnelle Läufer gewesen sein könnten.121Hutchinson, J. R. & Garcia, M. (2002). Tyrannosaurus was not a fast runner. Nature, 415(6875), 1018–1021. doi:10.1038/4151018a
 Thomas R. Holtz jr. Thomas R. Holtz jr. |
T. rex besaß allerdings mehrere anatomische Merkmale, die auch ihm eine effiziente Fortbewegung begünstigten. Dazu gehören relativ lange Unterschenkel und Füße im Vergleich zum Oberschenkelknochen sowie ein besonders stabil gebauter Mittelfuß. Dabei war der dritte Mittelfußknochen zwischen den beiden anderen eingeklemmt, wodurch eine Struktur entstand, die als Arctometatarsus bezeichnet wird und von Thomas R. Holtz jr. erstmalig als solche an T. rex erkannt wurde. Diese Konstruktion hat wahrscheinlich dazu beigetragen, die Kräfte beim Laufen effizient vom Fuß auf das Bein zu übertragen.122Holtz, T. R. (1996). Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda). Journal of Paleontology, 70(3), 536–538. doi:10.1017/S0022336000038506123Benton, M. (2014). Vertebrate palaeontology. John Wiley & Sons, 193. |
Dies machte T. rex zu einem effizienten Langstreckenläufer: 2020 haben Forscher die Beinproportionen, die Körpermasse und die Bewegungsweise von mehr als siebzig Theropodenarten verglichen. Dabei zeigte sich, dass lange Beine bei kleineren Theropoden vor allem einer hohen Geschwindigkeit dienten, während sie bei sehr großen Arten eher dazu beitrugen, beim eher gemächichen Gehen Energie zu sparen, was das Durchstreifen sehr großer Reviere ermöglichte.124Dececchi, T. A. et al. (2020). The fast and the frugal – divergent locomotory strategies drive limb lengthening in theropod dinosaurs. PLoS ONE, 15(5), e0223698. doi:10.1371/journal.pone.0223698 Eine Studie aus dem Jahr 2021 am Exemplar „Trix“ berechnete die bevorzugte Gehgeschwindigkeit von T. rex auf etwa 1,28 Meter pro Sekunde, also rund 4,6 km/h. Bei dieser Geschwindigkeit verbrauchen Tiere besonders wenig Energie, weil ihre Bewegungen im Rhythmus der natürlichen Schwingungen des Körpers erfolgen. Für Dinosaurier spielte dabei offenbar auch der Schwanz eine wichtige Rolle. Während des Gehens bewegte er sich leicht auf und ab, wobei Bänder Energie speicherten und wieder freisetzten.125van Bijlert, P. A. et al. (2021). Natural frequency method – estimating the preferred walking speed of Tyrannosaurus rex based on tail natural frequency. Royal Society Open Science, 8(4), 201441. doi:10.1098/rsos.201441
Dabei blieb T. rex trotzdem sehr beweglich: er konnte sich deutlich schneller um die eigene Achse drehen als andere Theropoden und auch seinen Lauf sehr plötzlich beschleunigen. Möglicherweise war er sogar in der Lage, sich während der Jagd auf einem Fuß zu drehen, während das andere Bein nach vorn schwang, ähnlich einer Pirouette.126Cotton, J. R. et al. (2019). Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods. PeerJ, 7, e6432. doi:10.7717/peerj.6432 Die maximale Laufgeschwindigkeit eines ausgewachsenen T. rex wird heute auf zwischen mindestens 15 bis maximal 30km/h beziffert,127Hirt, M. R. et al. (2017). A general scaling law reveals why the largest animals are not the fastest. Nature Ecology & Evolution, 1(8), 1116–1122. doi:10.1038/s41559-017-0241-4 wobei die höheren Werte aber aufgrund der enormen Belastung für Muskulatur, Gelenke und Knochen und auch einem extremen Verletzugsrisiko bei Stürzen als eher unwahrscheinlich angesehen werden.128Sellers, W. I. et al. (2017). Investigating the running abilities of Tyrannosaurus rex using stress-constrained multibody dynamic analysis. PeerJ, 5, e3420. doi:10.7717/peerj.3420 Jedoch gilt dies nur, wenn man eine relativ weite Schrittlänge veranschlagt.
Aufschluss über die tatsächliche Schrittlänge können uns fossile Fußabdrücke, sogenannte Ichnofossilien geben. Bislang wurden aber nur wenige solcher Fährten von T. rex beschrieben. Die erste wurde 1983 von Charles Pillmore auf der Philmont Scout Ranch entdeckt. Zunächst hielt man die Spur für die eines Hadrosauriers. Spätere Untersuchungen zeigten jedoch, dass sie zu einem großen Theropoden gehören mussten. Da T. rex wohl der einzige Großtheropode in seinem Lebensraum war, ist es nur wahrscheinlich, dass er der Verursacher war. 1994 beschrieben Martin Lockley und Adrian Hunt diese Spur unter dem Ichnogenus Tyrannosauripus pillmorei. Die Spur entstand in einem damals bewachsenen, schlammigen Feuchtgebiet und misst 83 Zentimeter Länge sowie 71 Zentimeter Breite.129Lockley, M. G. & Hunt, A. P. (1994). A track of the giant theropod dinosaur Tyrannosaurus from close to the Cretaceous/Tertiary boundary, northern New Mexico. Ichnos, 3(3), 213–218. doi:10.1080/10420949409386390 Eine zweite mögliche Tyrannosaurus-Spur wurde 2007 aus der Hell Creek Formation in Montana gemeldet. Diese Spur ist mit 72 Zentimetern etwas kürzer als das von Lockley und Hunt beschriebene Exemplar.130Manning, P. L., Ott, C. & Falkingham, P. L. (2009). The first tyrannosaurid track from the Hell Creek Formation (Late Cretaceous), Montana, U.S.A. PALAIOS, 23(10), 645–647. doi:10.2110/palo.2008.p08-030r Allerdings verraten solche einzelnen Abdrücke natürlich noch nichts über die Geschwindigkeit des Tieres, das sie einst hinterlließ. Um Rückschlüsse über die Geschwindigkeit zu gewinnen, bräuchte es eine ganze Fährte, also mehrere Fußabdrücke hintereinander.

So eine Fährtenreihe ist inzwischen aber aus Glenrock in Wyoming bekannt, also einem Teil der Lance Formation. Die Spuren wurden 2016 beschrieben und entweder von einem jegendlichen T. rex oder Nanotyrannus lancensis verursacht. Anhand der Maße und der Abstände der Spuren schätzten Forscher die Gehgeschwindigkeit des Tieres auf etwa 4,5 bis 8 Kilometer pro Stunde. Die Hüfthöhe wurde auf etwa 1,56 bis 2,06 Meter geschätzt.131Smith, S. D., Persons, W. S. & Xing, L. (2016). A Tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming. Cretaceous Research, 61(1), 1–4. doi:10.1016/j.cretres.2015.12.020 Eine Folgestudie aus dem Jahr 2017 erhöhte diese Geschwindigkeits-Schätzungen um weitere 50 bis 80 %, was dann etwa 7 bis 14 Kilometern pro Stunde entsprochen hätte. Noch zwei weitere fossile Fußspuren und Fährten, die ebenfalls Tyrannosauripus zugeordnet werden, stammen außerdem aus New Mexico und Wyoming. Diese Zuordnung basiert vor allem auf dem stratigraphischen Alter der Gesteinsschichten, in denen die Fossilien gefunden wurden. Zuletzt wurden zwei sehr spannende Fährten aus Ludlow in Colorado und Cimarron in New Mexico beschrieben. Sie wurden offenbar von einem großen Tyrannosauriden verursacht, der aus einer liegenden Position aufgestanden ist. Dabei hat sich das Tier wohl auf seine Ellenbogen und die Ballen der Füße aufgestützt.132Caneer, T., Molkestad, T. & Lucas, S. G. (2021). Tracks in the Upper Cretaceous of the Raton Basin possibly show tyrannosaurid rising from a prone position. New Mexico Museum of Natural History and Science, 29–37.
Neue Berechnungen, die auch unter Einbeziehung der Daten von den Fährten aangestellt wurden, konnten zeigen, dass sich der Fuß von T. rex beim Laufen eher wie der eines Vogels verhielt. Seine Schritte waren vergleichsweise kurz, dafür setzte er sie schneller hintereinander. Die so berechneten Höchstgeschwindigkeiten liegen für ausgewachsene Exemplare im Bereich von etwa 18 bis 25 Kilometern pro Stunde und stützen damit vorherige Daten. Somit steht relativ klar fest, dass erwachsene Exemplare wohl keine schnellen Sprinter, sondern eher ausdauernde, kraftvolle Läufer waren – ganz im Gegensatz zu den Jungtieren, die wohl mindestens 40km/h erreichen konnten! Ein ausgewachsener T. rex war allerdings immer noch so schnell wie ein durchschnittlicher Fahrradfahrer. Dieser Sachverhalt wird auch in meiner Romanreihe so dargestellt. 133Boeye, A. T. et al. (2026). Evidence of bird-like foot function in Tyrannosaurus. Royal Society Open Science, 13(2), 252139. doi:10.1002/10.1098/rsos.252139
Jagdverhalten
 Jack R. Horner Jack R. Horner |
Trotzdem hält sich die Vorstellung hartnäckig , dass T. rex vor allem ein langsamer Aasfresser gewesen sei, der zu träge war, um aktiv zu jagen. Die Diskussion darüber begann bereits 1917 mit einer Studie von Lawrence M. Lambe, der argumentierte, dass die Zähne von Gorgosaurus kaum Abnutzungsspuren zeigten, wie sie bei der Jagd hätten auftreten müssten.134Lambe, L. B. (1917). The Cretaceous theropodous dinosaur Gorgosaurus. Memoirs of the Geological Survey of Canada, 100, 1–84. doi:10.4095/101672 Damals war jedoch noch nicht bekannt, dass Theropoden ihre Zähne ständig ersetzten. Besonders in den 1990er Jahren bekam die Aasfresser-Hypothese mit Jack R. Horner ein prominentes Gesicht. |
Horner führte mehrere Punkte an, die für eine nichtjagende Lebensweise sprächen: Die Arme seien im Vergleich zu anderen Raubtieren extrem klein und hätten nicht genug Kraft, um Beute festzuhalten. Außerdem besaß T. rex außergewöhnlich große Riechkolben und Riechnerven. Dies deutet auf einen sehr ausgeprägten Geruchssinn hin, mit dem Kadaver über große Entfernungen aufgespürt werden konnten, ähnlich wie bei heutigen Geiern. Seine kräftigen Zähne konnten Knochen zerquetschen und so auch nährstoffreiches Knochenmark für die Verdauung nutzbar machen. Außerdem sei er viel langsamer als seine potenziellen Beutetiere gewesen.135Horner, J. R. (1994). Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger). The Paleontological Society Special Publication, 7, 157–164. doi:10.1017/S2475262200009497 Mit vielen dieser Punkte schien Horner auch erst einmal recht zu behalten: Tatsächlich zeigte eine Studie, dass T. rex unter 21 untersuchten Dinosauriern den am zweitstärksten entwickelten Geruchssinn besaß, übertroffen nur von seinem Verwandten Gorgosaurus libratus.136Zelenitsky, D. K., Therrien, F. & Kobayashi, Y. (2008). Olfactory acuity in theropods. Palaeobiological and evolutionary implications. Proceedings of the Royal Society B: Biological Sciences, 276(1657), 667–673. doi:10.1098/rspb.2008.1075 Außerdem konnten Forscher tatsächlich Knochenfragmente in Koprolithen nachweisen, die vermutlich von Tyrannosauriern stammen. Gleichzeitig wiesen sie jedoch darauf hin, dass die Zähne nicht so spezialisiert zum Knochenzermahlen waren wie bei Hyänen.137Chin, K., Tokaryk, T. T., Erickson, G. M. & Calk, L. C. (1998). A king-sized theropod coprolite. Nature, 393(6686), 680–682. doi:10.1038/31461
Andere Paläontologen wie Robert T. Bakker, James Farlow und Thomas R. Holtz Jr. hielten dagegen, dass viele heutige Räuber wie Wölfe, Hyänen oder Sekretärvögel ihre Vordergliedmaßen kaum zum Töten ihrer Beute einsetzen.138Farlow, J. O. & Holtz, T. R. (2002). The Fossil Record of Predation in Dinosaurs. The Paleontological Society Papers, 8, 251–266. doi:10.1017/S108933260000111X139Holtz, T. R. (2008). A critical re-appraisal of the obligate scavenging hypothesis for Tyrannosaurus rex and other tyrant dinosaurs. In Larson, P. L. & Carpenter, K. (Hgg.), Tyrannosaurus Rex, the Tyrant King. Indiana University Press, 371–394. Außerdem sei er vielleicht nicht so schnell wie heutige Raubtiere gewesen, doch hätte selbst das durch biomechanische Studien ermittelte Geschwindigkeitsspektrum allemal dazu ausgereicht, Hadrosaurier oder Ceratopsier auf kurzen Strecken einzuholen.
Eine Studie aus dem Jahr 2012 kam zu dem Ergebnis, dass T. rex den stärksten Biss aller bekannten landlebenden Tiere besaß. Ausgewachsene Individuen könnten demnach mit den hinteren Zähnen eine Kraft von etwa 35.000 bis 57.000 Newton erzeugt haben.140Bates, K. T. & Falkingham, P. L. (2012). Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics. Biology Letters, 8(4), 660–664. doi:10.1098/rsbl.2012.0056 Andere Schätzungen fielen sogar noch höher aus und landeten bei 183.000 bis 235.000 Newton.141Meers, M. B. (2003). Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior. Historical Biology, 16(1), 1–12. doi:10.1080/0891296021000050755 Außerdem wurde berechnet, dass T. rex sein Maul in einem weiten Winkel vin bis zu etwa 80 Grad öffnen konnte, was eine wichtige Anpassung für die enorme Beißkraft darstellt.142Lautenschlager, S. (2015). Estimating cranial musculoskeletal constraints in theropod dinosaurs. Royal Society Open Science, 2(11), 150495. doi:10.1098/rsos.150495 Aufgrund der flachen Gelenke der Halswirbel war der Schädel selbst wohl weniger beweglich als bei Allosauriern. Wahrscheinlich konnte T. rex seine Beute aber durch seitliche Kopfbewegungen effizient zerreißen, ganz ähnlich wie heutige Krokodile und Alligatoren.143Snively, E., Cotton, J. R., Ridgely, R. & Witmer, L. M. (2013). Multibody dynamics model of head and neck function in Allosaurus (Dinosauria, Theropoda). Palaeontologia Electronica, 16(2), 338. doi:10.26879/338
Auch andere Hinweise sprechen deutlich für aktives Jagdverhalten. Die Augenhöhlen von Tyrannosauriern waren nach vorne gerichtet, wodurch stereoskopisches Sehen mit guter Tiefenwahrnehmung möglich war. Für einen reinen Aasfresser wäre eine solche räumliche Wahrnehmung kaum nötig gewesen.144Stevens, K. A. (2006). Binocular vision in theropod dinosaurs. Journal of Vertebrate Paleontology, 26(2), 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2145Jaffe, E. (2006). Sight for ‚Saur Eyes. T. rex vision was among nature’s best. Science News, 170(1), 3–4. doi:10.2307/4017288
Den wohl schärfsten Widerspruch zur Aasfresser-Hypothese stelllt aber ein Edmontosaurus annectens-Skelett aus Montana dar. Dessen Schwanzwirbel zeigen verheilte Bissverletzungen eines Tyrannosauriers. Da der Hadrosaurier den Biss nachweislich überlebte, muss er einem Angriff entkommen sein.146Carpenter, K. (1998). Evidence of predatory behavior by theropod dinosaurs. Gaia, 15, 135–144. Auch zwischen Triceratops und Tyrannosaurus scheint es gewaltsame Begegnungen gegeben zu haben. Teilweise verheilte Zahnspuren eines Tyrannosauriers auf Horn und Nackenschild eines Triceratops zeigen, dass auch dieses Tier den Angriff überlebte.147Happ, J. & Carpenter, K. (2008). An analysis of predator–prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. In Carpenter, K. & Larson, P. E. (Hgg.), Tyrannosaurus rex, the Tyrant King. Indiana University Press, Bloomington, 355–368. Untersuchungen von Hadrosaurier-Wirbeln aus der Hell Creek Formation, die von mutmaßlich jugendlichen Tyrannosauriern durchbohrt wurden, zeigen zudem, dass selbst junge Tiere bereits tödliche Jäger gewesen sein dürften.148Peterson, J. E. & Daus, K. N. (2019). Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends. PeerJ, 7, e6573. doi:10.7717/peerj.6573 Auch Verletzungsspuren an T. rex-Exemplaren selbst deuten auf ein Leben hin, in dem die Tiere häufig enormen Belastungen ausgesetzt waren, auch durch Kämpfe mit ihren Artgenossen.149Tanke, D. H. & Currie, P. J. (1998). Head-biting behavior in theropod dinosaurs. Paleopathological evidence. Gaia, 15, 167–184. Eine sehr authentische Jagdszene wird in Prehistoric Planet (2023) gezeigt:
1992 schlug William Abler vor, T. rex könnte auch giftigen beziehungsweise hochinfektiösen Speichel besessen haben. Er argumentierte, die Bakterien aus den Zwischenräumen zwischen den Zahnzacken hätten einmal gebissenen Opfern ebenfalls über kurz oder lang den Tod bereitet. Ähnliches nahm man damals auch bei Komodowaranen an.150Abler, W. L. (1992). The serrated teeth of tyrannosaurid dinosaurs, and biting structures in other animals. Paleobiology, 18(2), 161–183. doi:10.1017/S0094837300013956 Diese Annahme konnte bei Waranen aber inzwischen widerlegt werden. Ihre Beute stirbt nach einem Biss vor allem an einem hochwirksamen Gift.151Goldstein, E. J. C.; Tyrrell, K. L.; Citron, D. M.; Cox, C. R.; Recchio, I. M.; Okimoto, B.; Bryja, J.; Fry, B. G. (June 1, 2013). „Anaerobic and aerobic bacteriology of the saliva and gingiva from 16 captive Komodo dragons (Varanus komodoensis): new implications for the ‚bacteria as venom‘ model“. Journal of Zoo and Wildlife Medicine. 44 (2): 262–272. doi:10.1638/2012-0022R.1. PMID 23805543. Da für bakterielle Infektionen als speziealisierte Tötungsmethode nun bei keinem Tier belegt sind, kann dies auch bei T. rex ausgeschlossen werden.
Heute gehen die meisten Forscher jedoch davon aus, dass T. rex aktiv jagte, aber opportunistisch auch Aas fraß oder Beute anderer Räuber stahl, genauso wie es heutige Großraubtiere tun.152Farlow, J. O. & Holtz, T. R. Jr. (2002). The fossil record of predation in dinosaurs. In Kowalewski, M. & Kelley, P. H. (Hgg.), The Fossil Record of Predation. The Paleontological Society Papers, 8, 251–266. Als mit Abstand größter Fleischfresser seiner Umwelt war T. rex wahrscheinlich ein Spitzenprädator, der Hadrosaurier, gepanzerte Pflanzenfresser wie Ceratopsier und Ankylosaurier und möglicherweise sogar Sauropoden wie Alamosaurus jagte.153Black, R. (2012). When Tyrannosaurus Chomped Sauropods. Smithsonian Magazine, 25, 469. doi:10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2 Untersuchungen von Kalziumisotopen im Zahnschmelz deuten zudem darauf hin, dass T. rex gelegentlich auch Kadaver von Meeresreptilien oder Fischen fraß, die an die Küsten des Western Interior Seaway gespült worden waren.154Martin, J. E., Hassler, A., Montagnac, G., Therrien, F. & Balter, V. (2022). The stability of dinosaur communities before the Cretaceous–Paleogene (K–Pg) boundary. A perspective from southern Alberta using calcium isotopes as a dietary proxy. Geological Society of America Bulletin, 134(9–10), 2548–2560. doi:10.1130/B36222.1

Quellenangaben:
- 1Dingus, L. & Norell, M. (2010). Barnum Brown – The man who discovered Tyrannosaurus rex. University of California Press, 90, 124.
- 2Osborn, H. F. (1905). Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the AMNH, 21(14), 259–265.
- 3Dingus, L. & Norell, M. (2010). Barnum Brown – The man who discovered Tyrannosaurus rex. University of California Press, 90, 124.
- 4Osborn, H. F. & Brown, B. (1906). Tyrannosaurus, Upper Cretaceous carnivorous dinosaur. Bulletin of the AMNH, 22(16), 281–296.
- 5Breithaupt, B. H. et al. (2006). Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West. New Mexico Museum of Natural History and Science Bulletin, 35, 258.
- 6Breithaupt, B. H. et al. (2005). In celebration of 100 years of Tyrannosaurus rex – Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the earliest discoveries of Tyrannosaurus rex in the West. Abstracts with Programs, 37(7), 406.
- 7Breithaupt, B. H. et al. (2005). In celebration of 100 years of Tyrannosaurus rex – Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the earliest discoveries of Tyrannosaurus rex in the West. Abstracts with Programs, 37(7), 406.
- 8Hatcher, J. B. (1907). The Ceratopsia. Monographs of the United States Geological Survey, 49, 113–114.
- 9Osborn, H. F. (1917). Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bulletin of the American Museum of Natural History, 35(43), 733–771.
- 10Larson, N. L. (2008). One hundred years of Tyrannosaurus rex – the skeletons. In Larson, P. & Carpenter, K. (Hgg.), Tyrannosaurus rex, the Tyrant King. Indiana University Press, 1–55.
- 11Brochu, C. R. (2003). Osteology of Tyrannosaurus rex – insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Society of Vertebrate Paleontology Memoirs, 7, 1–138. doi:10.2307/3889334
- 12Larson, N. L. (2008). One hundred years of Tyrannosaurus rex – the skeletons. In Larson, P. & Carpenter, K. (Hgg.), Tyrannosaurus rex, the Tyrant King. Indiana University Press, 1–55.
- 13Fiffer, S. (2000). Tyrannosaurus Sue. W. H. Freeman and Company, New York. 195-212.
- 14Escobedo, T. (2014). When the feds seized a T. rex. CNN.
- 15Browne, M. (1996). Fossil Dealer, Target of Federal Prosecutors, Begins Jail Term. New York Times.
- 16Regalado, A. (1995). Fossil dealers innocent – and guilty. Science, 267(5205).
- 17Small, Z. (2020). T. rex Skeleton Brings $31.8 Million at Christie’s Auction. The New York Times.
- 18Hutchinson, J. R. et al. (2011). A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth. PLOS ONE, 6(10), e26037. doi:10.1371/journal.pone.0026037
- 19Small, Z. (2020). T. rex Skeleton Brings $31.8 Million at Christie’s Auction. The New York Times.
- 20Greshko, M. (2022). Stan the T. rex found! World’s most expensive fossil finds home in a new museum. National Geographic.
- 21Martin, N. (2025). Exclusive. New Natural History Museum Abu Dhabi features two fighting T. rexes. National Geographic Society.
- 22Boeye, A. T. et al. (2026). Evidence of bird-like foot function in Tyrannosaurus. Royal Society Open Science, 13(2). doi:10.1098/rsos.252139
- 23Tanke, D. H. & Currie, P. J. (1998). Head-biting behavior in theropod dinosaurs. Paleopathological evidence. Gaia, 15, 167–184.
- 24Larson, P. L. & Carpenter, K. (2008). Tyrannosaurus rex, the Tyrant King. Life of the Past. Indiana University Press.
- 25Crosslin, R., Fortney, M. & Dinosphere Exhibit Development Team (2004). Resource Materials. Bucky-Tyrannosaurus rex. Dinosphere: A 3–5 Unit of Study.
- 26Larson, P. The First Recognized Furcula for Tyrannosaurus rex. Black Hills Institute of Geological Research.
- 27Larson, P. L. (2013). The case for Nanotyrannus. In Parrish, J. M., Molnar, R. A., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. University of Indiana Press, 15–53.
- 28Hutchinson, J. R. et al. (2011). A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth. PLOS ONE, 6(10), e26037. doi:10.1371/journal.pone.0026037
- 29Zanno, L. E. & Napoli, J. G. (2025). Nanotyrannus and Tyrannosaurus coexisted at the close of the Cretaceous. Nature, 648(8093), 357–367. doi:10.1038/s41586-025-09801-6
- 30Woodward, H. N., Myhrvold, N. P. & Horner, J. R. (2026). Prolonged growth and extended subadult development in the Tyrannosaurus rex species complex revealed by expanded histological sampling and statistical modeling. PeerJ, 14, e20469. doi:10.7717/peerj.20469
- 31Cullen, T. M. et al. (2020). Osteohistological analyses reveal diverse strategies of theropod dinosaur body-size evolution. Proceedings of the Royal Society B: Biological Sciences, 287(1939), 20202258. doi:10.1098/rspb.2020.2258
- 32Erickson, G. M. et al. (2006). Tyrannosaur life tables – an example of nonavian dinosaur population biology. Science, 313(5784), 213–217. doi:10.1126/science.1125721
- 33Farlow, J. O. (1989). Paleobiology of the Dinosaurs. Geological Society of America.
- 34Horner, J. R. & Makela, R. (1979). Nest of juveniles provides evidence of family structure among dinosaurs. Nature, 282(5736), 296–298. doi:10.1038/282296a0
- 35Norell, M. A. et al. (1995). A nesting dinosaur. Nature, 378(6559), 774–776. doi:10.1038/378774a0
- 36Watanabe, M. E. (2009). Evolving Ideas on the Origins of Parental Care. BioScience, 59(3), 272. doi:10.1525/bio.2009.59.3.17
- 37Horner, J. R. & Padian, K. (2004). Age and growth dynamics of Tyrannosaurus rex. Proceedings: Biological Sciences, 271(1551), 1875–1880. doi:10.1098/rspb.2004.2829
- 38Erickson, G. M. et al. (2006). Tyrannosaur life tables – an example of nonavian dinosaur population biology. Science, 313(5784), 213–217. doi:10.1126/science.1125721
- 39Hutchinson, J. R. et al. (2011). A computational analysis of limb and body dimensions in Tyrannosaurus rex with implications for locomotion, ontogeny, and growth. PLoS ONE, 6(10), e26037. doi:10.1371/journal.pone.0026037
- 40Schweitzer, M. H. et al. (2005). Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. Science, 308(5727), 1456–1460. doi:10.1126/science.1112158
- 41Lee, A. H. & Werning, S. (2008). Sexual maturity in growing dinosaurs does not fit reptilian growth models. Proceedings of the National Academy of Sciences, 105(2), 582–587. doi:10.1073/pnas.0708903105
- 42Schweitzer, M. H. et al. (2016). Chemistry supports the identification of gender-specific reproductive tissue in Tyrannosaurus rex. Scientific Reports, 6, 23099. doi:10.1038/srep23099
- 43Woodward, H. N. et al. (2026). Prolonged growth and extended subadult development in the Tyrannosaurus rex species complex revealed by expanded histological sampling and statistical modeling. PeerJ, 14, e20469. doi:10.7717/peerj.20469
- 44Zanno, L. E. & Napoli, J. G. (2025). Nanotyrannus and Tyrannosaurus coexisted at the close of the Cretaceous. Nature. doi:10.1038/s41586-025-09801-6
- 45Paul, G. S. (2008). Chapter 18 – The extreme life style and habits of the gigantic tyrannosaurid superpredators of the Cretaceous North America and Asia. In Larson, P. L. & Carpenter, K. (Hgg.), Tyrannosaurus, the Tyrant King. Indiana University Press, 307–345.
- 46Holtz, T. R. Jr. (2013). The life and times of Tyrannosaurus rex, with Dr. Thomas Holtz. Burke Museum of Natural History and Culture.
- 47Leidy, J. (1865). Memoir on the extinct reptiles of the Cretaceous formations of the United States. Cretaceous reptiles of the United States. Smithsonian Contributions to Knowledge, 14, 1–4. doi:10.5962/bhl.title.39830
- 48Newman, B. H. (1970). Stance and gait in the flesh-eating Tyrannosaurus. Biological Journal of the Linnean Society, 2(2), 119–123. doi:10.1111/j.1095-8312.1970.tb01707.x
- 49Padian, K. (2022). Why tyrannosaurid forelimbs were so short. An integrative hypothesis. Acta Palaeontologica Polonica, 67(1), 63–76.
- 50Stevens, K. A., Larson, P., Willis, E. D. & Anderson, A. (2008). Rex, sit. Digital modeling of Tyrannosaurus rex at rest. In Larson, P. & Carpenter, K. (Hgg.), Tyrannosaurus rex, the tyrant king. Indiana University Press, 192–203.
- 51Lambe, L. M. (1914). On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. Ottawa Naturalist, 27, 129–135.
- 52Horner, J. R. & Lessem, D. (1993). The complete T. rex. Simon & Schuster, New York City.
- 53Osborn, H. F. & Brown, B. (1906). Tyrannosaurus, Upper Cretaceous carnivorous dinosaur. Bulletin of the AMNH, 22(16), 281–296.
- 54Carpenter, K. & Smith, M. (2001). Forelimb Osteology and Biomechanics of Tyrannosaurus rex. In Tanke, D. H. & Carpenter, K. (Hgg.), Mesozoic vertebrate life. Indiana University Press, Bloomington, 90–116.
- 55Pickrell, J. (2017). T. Rex’s Tiny Arms May Have Been Vicious Weapons. National Geographic.
- 56Padian, K. (2022). Why tyrannosaurid forelimbs were so short. An integrative hypothesis. Acta Palaeontologica Polonica, 67(1), 63–76.
- 57Ostrom, J. H. (1969). Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Bulletin of the Peabody Museum of Natural History, 30.
- 58Bakker, R. T. (1972). Anatomical and ecological evidence of endothermy in dinosaurs. Nature, 238(5359), 81–85. doi:10.1038/238081a0
- 59Bakker, R. T. (1986). The Dinosaur Heresies. Kensington Publishing, New York, 241.
- 60Horner, J. R. & Padian, K. (2004). Age and growth dynamics of Tyrannosaurus rex. Proceedings: Biological Sciences, 271(1551), 1875–1880. doi:10.1098/rspb.2004.2829
- 61Barrick, R. E. & Showers, W. J. (1994). Thermophysiology of Tyrannosaurus rex. Evidence from Oxygen Isotopes. Science, 265(5169), 222–224. doi:10.1126/science.265.5169.222
- 62Wiemann, J. et al. (2022). Fossil biomolecules reveal an avian metabolism in the ancestral dinosaur. Nature, 606(7914), 522–526. doi:10.1038/s41586-022-04770-6
- 63Holliday, C. M. et al. (2019). The Frontoparietal Fossa and Dorsotemporal Fenestra of Archosaurs and Their Significance for Interpretations of Vascular and Muscular Anatomy in Dinosaurs. The Anatomical Record, 303(4), 1060–1074. doi:10.1002/ar.24218
- 64Paladino, F. V., Spotila, J. R. & Dodson, P. (1999). A blueprint for giants. Modeling the physiology of large dinosaurs. In Farlow, J. O. & Brett-Surman, M. K. (Hgg.), The Complete Dinosaur. Indiana University Press, Bloomington, 491–504.
- 65Chinsamy, A. & Hillenius, W. J. (2004). Physiology of nonavian dinosaurs. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The dinosauria. University of California Press, Berkeley, 643–659.
- 66Seymour, R. S. (2013). Maximal Aerobic and Anaerobic Power Generation in Large Crocodiles versus Mammals. Implications for Dinosaur Gigantothermy. PLOS ONE, 8(7), e69361. doi:10.1371/journal.pone.0069361
- 67Xing, X. et al. (2004). Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature, 431(7009), 680–684. doi:10.1038/nature02855
- 68Xing, X. et al. (2012). A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature, 484(7392), 92–95. doi:10.1038/nature10906
- 69Bell, P. R. et al. (2017). Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution. Biology Letters, 13(6), 20170092. doi:10.1098/rsbl.2017.0092
- 70Reisz, R. R. & Larson, D. (2016). Dental anatomy and skull length to tooth size ratios support the hypothesis that theropod dinosaurs had lips. 4th Annual Meeting, 2016, Canadian Society of Vertebrate Palaeontology.
- 71Carr, T. D. et al. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7, 44942. doi:10.1038/srep44942
- 72Milinkovitch, M. et al. (2013). Crocodile head scales are not developmental units but emerge from physical cracking. Science, 339(6115), 78–81. doi:10.1126/science.1226265
- 73Cullen, T. M. et al. (2023). Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology. Science, 379(6639), 1348–1352. doi:10.1126/science.abo7877
- 74Schweitzer, M. H. et al. (2005). Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science, 307(5717), 1952–1955. doi:10.1126/science.1108397
- 75Anné, J. et al. (2023). Independent Evidence for the Preservation of Endogenous Bone Biochemistry in a Specimen of Tyrannosaurus rex. Biology, 12(2), 264. doi:10.3390/biology12020264
- 76Kaye, T. G. et al. (2008). Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms. PLOS ONE, 3(7), e2808. doi:10.1371/journal.pone.0002808
- 77San Antonio, J. D. et al. (2011). Dinosaur Peptides Suggest Mechanisms of Protein Survival. PLOS ONE, 6(6), e20381. doi:10.1371/journal.pone.0020381
- 78Peterson, J. E., Lenczewski, M. E. & Scherer, R. P. (2010). Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs. PLOS ONE, 5(10), e13334. doi:10.1371/journal.pone.0013334
- 79Carpenter, K. (1992). Variation in Tyrannosaurus rex. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Approaches and Perspectives. Cambridge University Press, Cambridge, 141–145.
- 80Larson, P. L. (1994). Tyrannosaurus sex. In Rosenberg, G. D. & Wolberg, D. L. (Hgg.), Dino Fest. The Paleontological Society Special Publications, 139–155.
- 81Erickson, G. M., Kristopher, L. A. & Larson, P. (2005). Androgynous rex. The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs. Zoology, 108(4), 277–286. doi:10.1016/j.zool.2005.08.001
- 82Brochu, C. R. (2003). Osteology of Tyrannosaurus rex. Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Society of Vertebrate Paleontology Memoirs, 7, 1–138. doi:10.2307/3889334
- 83Schweitzer, M. H., Wittmeyer, J. L. & Horner, J. R. (2005). Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. Science, 308(5727), 1456–1460. doi:10.1126/science.1112158
- 84Schweitzer, M. H. et al. (2007). Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? Bone, 40(4), 1152–1158. doi:10.1016/j.bone.2006.10.029
- 85Paul, G. S., Persons IV, W. S. & van Raalte, J. (2022). The Tyrant Lizard King, Queen and Emperor. Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus. Evolutionary Biology, 49(2), 156–179. doi:10.1007/s11692-022-09561-5
- 86Carr, T. D. et al. (2022). Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America. A Comment on „The Tyrant Lizard King, Queen and Emperor. Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus“. Evolutionary Biology, 49(3), 314–341. doi:10.1007/s11692-022-09573-1
- 87Paul, G. S. (2025). A presentation of the current data on the exceptionally diverse non-tyrannosaurid eutyrannosaur and Tyrannosaurini genera and species of western North America during the End Cretaceous North American Interchange. Mesozoic, 2(2), 085–138. doi:10.11646/mesozoic.2.2.1
- 88Lozinsky, R. P., Hunt, A. P., Wolberg, D. L. & Lucas, S. G. (1984). Late Cretaceous (Lancian) dinosaurs from the McRae Formation, Sierra County, New Mexico. New Mexico Geology, 6(4), 72–77. doi:10.58799/NMG-v6n4.72
- 89Gillette, D. D., Wolberg, D. L. & Hunt, A. P. (1986). Tyrannosaurus rex from the McRae Formation (Lancian, Upper Cretaceous), Elephant Butte reservoir, Sierra County, New Mexico. New Mexico Geological Society Guidebook, 37, 235–238. doi:10.56577/FFC-37.235
- 90Lehman, T. M. & Carpenter, K. (1990). A partial skeleton of the tyrannosaurid dinosaur Aublysodon from the Upper Cretaceous of New Mexico. Journal of Paleontology, 64(6), 1026–1032. doi:10.1017/S0022336000019843
- 91Carr, T. D. & Williamson, T. E. (2000). A review of Tyrannosauridae (Dinosauria: Coelurosauria) from New Mexico. Bulletin, 17. New Mexico Museum of Natural History and Science, 113–145.
- 92Sullivan, R. M. & Lucas, S. G. (2015). Cretaceous Vertebrates of New Mexico. New Mexico Museum of Natural History and Science Bulletin, 68.
- 93Dalman, S. G. et al. (2024). A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism. Scientific Reports, 13(1), 22124. doi:10.1038/s41598-023-47011-0
- 94Schantz, E. & Amato, J. M. (2024). Dating dinosaurs in the Laramide foreland. U-Pb geochronologic constraints on a stratigraphic section containing Alamosaurus and the postulated Tyrannosaurus mcraeensis in the Love Ranch Basin. New Mexico Geological Society of America Abstracts with Programs, 56, 9.
- 95Scherer, C. R. (2025). Multiple lines of evidence support anagenesis in Daspletosaurus and cladogenesis in derived tyrannosaurines. Cretaceous Research, 169, 106080. doi:10.1016/j.cretres.2025.106080
- 96Morrison, C. et al. (2025). Rise of the king. Gondwanan origins and evolution of megaraptoran dinosaurs. Royal Society Open Science, 12(5), 242238. doi:10.1098/rsos.242238
- 97Voris, J. T. et al. (2025). A new Mongolian tyrannosauroid and the evolution of Eutyrannosauria. Nature, 642(8069), 973–979. doi:10.1038/s41586-025-08964-6
- 98Carr, T. D. (2025). Observations on the skull of the type specimen of Tyrannosaurus rex Osborn, 1905. All Earth, 37(1), 2539638, 1–66. doi:10.1080/27669645.2025.2539638
- 99Stevens, K. A. (2006). Binocular vision in theropod dinosaurs. Journal of Vertebrate Paleontology, 26(2), 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2
- 100Jaffe, E. (2006). Sight for Saur Eyes. T. rex vision was among nature’s best. Science News, 170(1), 3–4. doi:10.2307/4017288
- 101Holtz, T. R. Jr. (2013). The Life and Times of Tyrannosaurus rex, with Dr. Thomas Holtz. Lecture, Burke Museum of Natural History and Culture, Seattle, WA.
- 102Carpenter, K. (2013). A Closer Look at the Scavenging versus Predation by Tyrannosaurus rex. In Parrish, M. J., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. Life of the Past. Indiana University Press, Bloomington, 265–278.
- 103Witmer, L. M. & Ridgely, R. C. (2009). New Insights into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior. The Anatomical Record, 292(9), 1266–1296. doi:10.1002/ar.20983
- 104Zelenitsky, D. K., Therrien, F. & Kobayashi, Y. (2008). Olfactory acuity in theropods. Palaeobiological and evolutionary implications. Proceedings of the Royal Society B: Biological Sciences, 276(1657), 667–673. doi:10.1098/rspb.2008.1075
- 105Carr, T. D. et al. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7, 44942. doi:10.1038/srep44942
- 106Kawabe, S. & Hattori, S. (2021). Complex neurovascular system in the dentary of Tyrannosaurus. Historical Biology, 34(7), 1137–1145. doi:10.1080/08912963.2021.1965137
- 107Bouabdellah, F., Lessner, E. & Benoit, J. (2022). The rostral neurovascular system of Tyrannosaurus rex. Palaeontologia Electronica, 25(1), 1–20. doi:10.26879/1178
- 108Brochu, C. A. (2000). A digitally-rendered endocast for Tyrannosaurus rex. Journal of Vertebrate Paleontology, 20(1), 1–6. doi:10.1671/0272-4634(2000)020[0001:ADREFT]2.0.CO;2
- 109Hurlburt, G. S., Ridgely, R. C. & Witmer, L. M. (2013). Relative size of brain and cerebrum in Tyrannosaurid dinosaurs. An analysis using brain-endocast quantitative relationships in extant alligators. In Parrish, M. J., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. Indiana University Press, 134–154.
- 110Brusatte, S. (2018). The Rise and Fall of the Dinosaurs. HarperCollins Publishers, New York, NY, 219.
- 111Herculano-Houzel, S. (2023). Theropod dinosaurs had primate-like numbers of telencephalic neurons. Journal of Comparative Neurology, 531(9), 962–974. doi:10.1002/cne.25453
- 112Caspar, K. R. et al. (2024). How smart was T. rex? Testing claims of exceptional cognition in dinosaurs and the application of neuron count estimates in palaeontological research. The Anatomical Record, 307(12), 3685–3716. doi:10.1002/ar.25459
- 113Currie, P. J. (1998). Possible evidence of gregarious behaviour in tyrannosaurids. Gaia, 15, 271–277.
- 114Roach, B. T. & Brinkman, D. T. (2007). A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. Bulletin of the Peabody Museum of Natural History, 48(1), 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2
- 115Collins, N. (2011). Tyrannosaurus rex hunted in packs. The Telegraph.
- 116McCrea, R. T. (2014). A Terror of Tyrannosaurs. The First Trackways of Tyrannosaurids and Evidence of Gregariousness and Pathology in Tyrannosauridae. PLOS ONE, 9(7), e103613. doi:10.1371/journal.pone.0103613
- 117Rothschild, B. M. (2013). Clawing Their Way to the Top. Tyrannosaurid Pathology and Lifestyle. In Parrish, M. J., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (Hgg.), Tyrannosaurid Paleobiology. Life of the Past. Indiana University Press, Bloomington, 211–222.
- 118Anné, J. et al. (2023). Unusual lesions seen in the caudals of the hadrosaur, Edmontosaurus annectens. The Anatomical Record, 306(3), 594–606. doi:10.1002/ar.25078
- 119Longrich, N. R. et al. (2010). Cannibalism in Tyrannosaurus rex. PLOS ONE, 5(10), e13419. doi:10.1371/journal.pone.0013419
- 120Hutchinson, J. R. (2004). Biomechanical modeling and sensitivity analysis of bipedal running ability. II. Extinct taxa. Journal of Morphology, 262(1), 441–461. doi:10.1002/jmor.10240
- 121Hutchinson, J. R. & Garcia, M. (2002). Tyrannosaurus was not a fast runner. Nature, 415(6875), 1018–1021. doi:10.1038/4151018a
- 122Holtz, T. R. (1996). Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda). Journal of Paleontology, 70(3), 536–538. doi:10.1017/S0022336000038506
- 123Benton, M. (2014). Vertebrate palaeontology. John Wiley & Sons, 193.
- 124Dececchi, T. A. et al. (2020). The fast and the frugal – divergent locomotory strategies drive limb lengthening in theropod dinosaurs. PLoS ONE, 15(5), e0223698. doi:10.1371/journal.pone.0223698
- 125van Bijlert, P. A. et al. (2021). Natural frequency method – estimating the preferred walking speed of Tyrannosaurus rex based on tail natural frequency. Royal Society Open Science, 8(4), 201441. doi:10.1098/rsos.201441
- 126Cotton, J. R. et al. (2019). Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods. PeerJ, 7, e6432. doi:10.7717/peerj.6432
- 127Hirt, M. R. et al. (2017). A general scaling law reveals why the largest animals are not the fastest. Nature Ecology & Evolution, 1(8), 1116–1122. doi:10.1038/s41559-017-0241-4
- 128Sellers, W. I. et al. (2017). Investigating the running abilities of Tyrannosaurus rex using stress-constrained multibody dynamic analysis. PeerJ, 5, e3420. doi:10.7717/peerj.3420
- 129Lockley, M. G. & Hunt, A. P. (1994). A track of the giant theropod dinosaur Tyrannosaurus from close to the Cretaceous/Tertiary boundary, northern New Mexico. Ichnos, 3(3), 213–218. doi:10.1080/10420949409386390
- 130Manning, P. L., Ott, C. & Falkingham, P. L. (2009). The first tyrannosaurid track from the Hell Creek Formation (Late Cretaceous), Montana, U.S.A. PALAIOS, 23(10), 645–647. doi:10.2110/palo.2008.p08-030r
- 131Smith, S. D., Persons, W. S. & Xing, L. (2016). A Tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming. Cretaceous Research, 61(1), 1–4. doi:10.1016/j.cretres.2015.12.020
- 132Caneer, T., Molkestad, T. & Lucas, S. G. (2021). Tracks in the Upper Cretaceous of the Raton Basin possibly show tyrannosaurid rising from a prone position. New Mexico Museum of Natural History and Science, 29–37.
- 133Boeye, A. T. et al. (2026). Evidence of bird-like foot function in Tyrannosaurus. Royal Society Open Science, 13(2), 252139. doi:10.1002/10.1098/rsos.252139
- 134Lambe, L. B. (1917). The Cretaceous theropodous dinosaur Gorgosaurus. Memoirs of the Geological Survey of Canada, 100, 1–84. doi:10.4095/101672
- 135Horner, J. R. (1994). Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger). The Paleontological Society Special Publication, 7, 157–164. doi:10.1017/S2475262200009497
- 136Zelenitsky, D. K., Therrien, F. & Kobayashi, Y. (2008). Olfactory acuity in theropods. Palaeobiological and evolutionary implications. Proceedings of the Royal Society B: Biological Sciences, 276(1657), 667–673. doi:10.1098/rspb.2008.1075
- 137Chin, K., Tokaryk, T. T., Erickson, G. M. & Calk, L. C. (1998). A king-sized theropod coprolite. Nature, 393(6686), 680–682. doi:10.1038/31461
- 138Farlow, J. O. & Holtz, T. R. (2002). The Fossil Record of Predation in Dinosaurs. The Paleontological Society Papers, 8, 251–266. doi:10.1017/S108933260000111X
- 139Holtz, T. R. (2008). A critical re-appraisal of the obligate scavenging hypothesis for Tyrannosaurus rex and other tyrant dinosaurs. In Larson, P. L. & Carpenter, K. (Hgg.), Tyrannosaurus Rex, the Tyrant King. Indiana University Press, 371–394.
- 140Bates, K. T. & Falkingham, P. L. (2012). Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics. Biology Letters, 8(4), 660–664. doi:10.1098/rsbl.2012.0056
- 141Meers, M. B. (2003). Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior. Historical Biology, 16(1), 1–12. doi:10.1080/0891296021000050755
- 142Lautenschlager, S. (2015). Estimating cranial musculoskeletal constraints in theropod dinosaurs. Royal Society Open Science, 2(11), 150495. doi:10.1098/rsos.150495
- 143Snively, E., Cotton, J. R., Ridgely, R. & Witmer, L. M. (2013). Multibody dynamics model of head and neck function in Allosaurus (Dinosauria, Theropoda). Palaeontologia Electronica, 16(2), 338. doi:10.26879/338
- 144Stevens, K. A. (2006). Binocular vision in theropod dinosaurs. Journal of Vertebrate Paleontology, 26(2), 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2
- 145Jaffe, E. (2006). Sight for ‚Saur Eyes. T. rex vision was among nature’s best. Science News, 170(1), 3–4. doi:10.2307/4017288
- 146Carpenter, K. (1998). Evidence of predatory behavior by theropod dinosaurs. Gaia, 15, 135–144.
- 147Happ, J. & Carpenter, K. (2008). An analysis of predator–prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. In Carpenter, K. & Larson, P. E. (Hgg.), Tyrannosaurus rex, the Tyrant King. Indiana University Press, Bloomington, 355–368.
- 148Peterson, J. E. & Daus, K. N. (2019). Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends. PeerJ, 7, e6573. doi:10.7717/peerj.6573
- 149Tanke, D. H. & Currie, P. J. (1998). Head-biting behavior in theropod dinosaurs. Paleopathological evidence. Gaia, 15, 167–184.
- 150Abler, W. L. (1992). The serrated teeth of tyrannosaurid dinosaurs, and biting structures in other animals. Paleobiology, 18(2), 161–183. doi:10.1017/S0094837300013956
- 151Goldstein, E. J. C.; Tyrrell, K. L.; Citron, D. M.; Cox, C. R.; Recchio, I. M.; Okimoto, B.; Bryja, J.; Fry, B. G. (June 1, 2013). „Anaerobic and aerobic bacteriology of the saliva and gingiva from 16 captive Komodo dragons (Varanus komodoensis): new implications for the ‚bacteria as venom‘ model“. Journal of Zoo and Wildlife Medicine. 44 (2): 262–272. doi:10.1638/2012-0022R.1. PMID 23805543.
- 152Farlow, J. O. & Holtz, T. R. Jr. (2002). The fossil record of predation in dinosaurs. In Kowalewski, M. & Kelley, P. H. (Hgg.), The Fossil Record of Predation. The Paleontological Society Papers, 8, 251–266.
- 153Black, R. (2012). When Tyrannosaurus Chomped Sauropods. Smithsonian Magazine, 25, 469. doi:10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2
- 154Martin, J. E., Hassler, A., Montagnac, G., Therrien, F. & Balter, V. (2022). The stability of dinosaur communities before the Cretaceous–Paleogene (K–Pg) boundary. A perspective from southern Alberta using calcium isotopes as a dietary proxy. Geological Society of America Bulletin, 134(9–10), 2548–2560. doi:10.1130/B36222.1
Hat dir dieser Beitrag gefallen?
Für noch mehr Lesestoff schau gerne in die Kategorie Meine Bücher. Alle meine Werke findest du auch in der Sidebar rechts. Dort gibt es auch einen Direktlink zu Amazon, wo du sie gleich bestellen kannst. ➡️
Wie kannst du Die Weißen Steine sonst noch unterstützen?
Klar, in erster Linie natürlich, wenn du dir eines meiner Bücher kaufst. Aber du kannst mir auch auf anderen Wegen eine große Freude machen:
- Teile diesen Beitrag auf deinen Social-Media-Kanälen!
- Schreibe mir eine nette Rezension auf Amazon!
- Kaufe hier auf der Website über Affiliate Links ein!
Ich kriege übrigens auch eine kleine Unterstützung, wenn du etwas völlig anderes kaufst. Es muss nicht unbedingt der hier beworbene Artikel sein! Jeder kleine Centbetrag hilft, diese Seite am Laufen zu erhalten.
 |
Zusammen mit meiner lieben Frau Sahar habe ich auch noch eine Menge mehr zu bieten! Sahar ist nämlich eine großartige Künstlerin und fertigt Gemälde, Lesezeichen, Stofftaschen und viele andere coole Artikel an. Auch mit tollen Urzeit-Motiven! Klick dich einfach mal durch die Beiträge in der Kategorie Produkte & Services. Bestimmt findest du da etwas nach deinem Geschmack. Oder auch ein schönes Geschenk für deine Lieben. |
Einen Grund, anderen eine Freude zu bereiten, findet man schließlich immer!
You could definitely see your enthusiasm within the article you write.
The world hopes for more passionate writers like you
who are not afraid to mention how they believe.
Always go after your heart.