
Name: Thescelosaurus neglectus („Vernachlässigte wunderbare Echse“)
Beschrieben: 1913 von Charles W. Gilmore
Ordnung: Ornithischia; Familie: Thescelosauridae
Länge: ♂ bis zu 4,3m, ♀ bis 3m
Gewicht: ♂ bis 300kg, ♀ bis 75kg
Ernährung: herbivor
Beschreibung:
Der flinke Thescelosaurus lebt in kleinen Familienclans von selten mehr als zehn Tieren, die von einem erwachsenen Männchen angeführt werden. Männchen werden fast dreimal so groß und schwer wie die Weibchen und haben eine gräulich-grün geperlte, ledrige Haut, die von einem hellvioletten Aalstrich geziert wird. Weibchen sind um einiges zierlicher und braun-gelb gestreift. Sie haben einen grünen Aalstrich auf dem Rücken und sind vor Feinden sehr viel besser getarnt als ihre größeren, männlichen Artgenossen. Die Jungtiere sind von einem hellbeigen Flaum bedeckt, der sich im Erwachsenenalter zu dicken, dunklen Filamentborsten verwächst, die dann nur noch ihren Rücken und die Flanken zieren. Sie dienen nicht mehr nur zur Wärmeisolation, sondern sowohl als Merkmal zur Arterkennung, bei den Männchen auch der Balz. Den Weibchen leisten sie auch als hochsensible Tastorgane gute Dienste, mit denen sie sich unter der Erde orientieren. Thescelosaurus lebt nämlich nicht nur auf den Farnprärien und in den Koniferenwäldern von Hell Creek, sondern auch zeitweilig in unterirdischen Bauen, wo die Weibchen ihre Jungen aufziehen.

Die Tiere sind vor allem tag- und dämmerungsaktiv und suchen bevorzugt am frühen Morgen und am späten Abend nach Nahrung. Die Nächte verbringen sie in ihrem Bau. Dort sind sie sicher vor den meisten Fressfeinden, und die labyrinthartigen Tunnel bieten ihnen Schutz selbst vor kleineren Räubern wie Pectinodon. Diese Bauten werden oft über mehrere Generationen hinweg genutzt und stetig erweitert, sodass sie ein weit verzweigtes Netzwerk unter der Erde bilden. Bei Gefahr stößt der wachhabende Thescelosaurus einen hellen Pfiff aus, der die anderen Herdenmitglieder warnt. Im Nu setzt sich dann der ganze Clan schnell in Bewegung und flieht. Selbst auf steilem Terrain bleibt er erstaunlich trittsicher und kann plötzliche Richtungswechsel mit Leichtigkeit ausführen. Junge Tiere trainieren ihre Wendigkeit spielerisch, indem sie einander jagen und dabei Hindernisse überspringen. Diese spielerischen Verfolgungsjagden helfen ihnen nicht nur, ihre Reflexe zu schärfen, sondern stärken auch den sozialen Zusammenhalt innerhalb des Clans.
Lebensweise:
Die Clans sind die meiste Zeit des Jahres an eigene Reviere gebunden, weshalb es nur sehr selten zu Auseinandersetzungen mit anderen Clans kommt. In der Paarungszeit schließen sich manchmal sogar zwei wandernde Clans zusammen, um Inzucht zu vermeiden. Bei so einem Zusammenschluss finden zunächst erbitterte Rangkämpfe zwischen den männlichen Alphatieren statt. Ähnlich aggressiv zeigen sich auch die Weibchen, wenn sie einen Brutplatz gefunden und für sich in Anspruch genommen haben. Bei innerartlichen Kämpfen kann es mitunter sehr rabiat zugehen, wenn die Tiere sich gegenseitig rammen, mit den Vorderfüßen nacheinander schlagen und mit den bekrallten Füßen heftige Tritte austeilen.

Meist ist Thescelosaurus aber ein eher friedliebender Dinosaurier, der sich nur selten aggressiv verhält. Obwohl er eher scheu ist, ist Thescelosaurus auch neugierig und verfügt über ausgezeichnete Instinkte. Allerdings werden Jungtiere, denen noch die Erfahrung der Älteren fehlt, wegen ihrer Unvorsichtigkeit oft zur Beute von strategisch vorgehenden Jägern wie Pectinodon. Gegen Raubtiere kann sich Thescelosaurus kaum zur Wehr setzen. Doch er kann eine Gefahr schon aus weiter Entfernung wahrnehmen, denn er verfügt über eine exzellente Witterung. Thescelosaurus gehört, auch wenn man ihn wegen seiner obskuren Lebensweise nur selten zu Gesicht bekommt, zu den häufigsten Dinosauriern in seinem Lebensraum.
Thescelosaurus in Die Weißen Steine:
 |
Band I:Im Kapitel „Dimension“ tritt eine Thescolosaurus-Herde auf. Es sind die ersten Dinosaurier, die John nach seiner Flucht aus der Höhle zu Gesicht bekommt. |
 |
Band II:Im Kapitel „Guter Stoff“ erscheint eine Thescelosaurus-Herde am Seeufer, wo die Mädchen gerade Hanf-Fasern verarbeiten, und macht sich über die Früchte ihrer Arbeit her. In „Gute Freundinnen“ können Sarah und Vanessa die scheuen Tiere allerdings vertreiben. |
 |
Band III:Im Kapitel „Game over“ begegnet Max einer Thescelosaurus-Herde. Auch in „Geheimnisse“ wird erwähnt, dass sie zu den Menschen immer weniger Scheu zeigen und ständig versuchen, ihnen die Vorräte zu stibitzen. |
 |
Band IV:Thescelosaurus tritt in diesem Buch nicht direkt auf. Es befinden sich aber mehrere Exemplare unter den Dinosaurier, die in „Der sterbende See“ am vulkanischen Giftgas erstickt sind. |
 |
Hier geht’s zurück zur Hauptseite Leben in der Urzeit! |

Wissenschaftliches zu Thescelosaurus:
 |
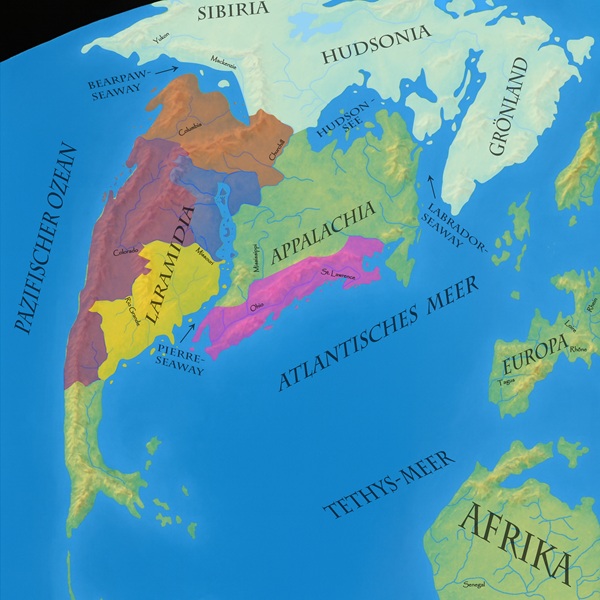
Thescelosaurus ist ein kleiner Ornithopode aus dem westlichen Nordamerika und zählt zu den häufihgsten dort gefunden Dinosauriern, auch wenn die Zuordnung mancher gefundener Exemplare nach wie vor unsicher ist. Sicher belegt ist er aber aus der Lance– sowie der Hell Creek Formation in den USA, auch in der kanadischen Scollard– und Frenchman Formation kam er sehr wahrscheinlich vor. Unsicher ist die Zuordnung bei Fossilien aus der Laramie-, Ferris-, Medicine Bow-, Almond- sowie der Willow Creek Formation. Fossilien aus der älteren Horseshoe Canyon Formation sowie der Prince Creek Formation in Alaska gehören wohl eher zu Parksosaurus.1Weishampel, D. B. et al. (2004). Dinosaur Distribution. In Weishampel, D. B., Osmólska, H. & Dodson, P. (Hrsg.), The Dinosauria. University of California Press, 517–606.2Brown, C. M. & Druckenmiller, P. (2011). Basal ornithopod (Dinosauria: Ornithischia) teeth from the Prince Creek Formation (early Maastrichtian) of Alaska. Canadian Journal of Earth Sciences, 48(9), 1342–1354. doi:10.1139/E11-017 Dennoch war Thescelosaurus im westlichen Nordamerika weit verbreitet und kam in allen küstennahen Lebensräumen im nördlichen Teil Laramidias vor. |
Forschungsgeschichte
Die ersten Fossilien von Thescelosaurus wurden während der sogenannten Bone Wars entdeckt, einer erbitterten Fehde zwischen den Paläontologen Edward D. Cope und Othniel C. Marsh. Seither sind viele weitere Skelette gefunden wurden, sodass wir den kompletten Körperbau dieses Tieres, seine Physiologie und auch Sinneseigenschaften und mögliche Lebensweise sehr gut erforschen können. Damit gehört Thescelosaurus zu den am besten bekannten, aber auch faszinierendsten Dinosauriern der späten Kreidezeit Nordamerikas.
 John B. Hatcher John B. Hatcher |
Im Juli 1891 entdeckte der Fossiliensammler John B.Hatcher, der damals für Marsh arbeitete, zusammen mit seinem Assistenten William H. Utterback ein nahezu vollständiges Skelett eines kleinen pflanzenfressenden Dinosauriers entlang des Doegie Creek im Niobrara County, Wyoming. |
Die Fossilien lagen in Gesteinen der Lance Formation. Das Skelett lag auf der linken Körperseite und war größtenteils noch in natürlicher anatomischer Verbindung erhalten, lediglich Kopf und Hals waren durch Erosion verloren gegangen. Der Fund wurde an das United States National Museum in Washington D.C. geschickt, das heute als National Museum of Natural History der Smithsonian Institution (USNM) bekannt ist. Marsh bekam den Fund jedoch niemals zu Gesicht. Das Skelett verblieb in einer unbeschrifteten Transportkiste in den Archivräumen des Museums und wurde über 20 Jahre lang nicht beachtet, was dem großen Umfang der Fossiliensammlung geschuldet war, die während der Bone Wars von Marsh zusammengetragen worden war.3Boyd, C. A. (2016). The „overlooked marvelous reptile“. Geo News, 43(1), 2–6.
Die „zweite“ Entdeckung und Benennung
 Charles W. Gilmore Charles W. Gilmore |
Im Jahre 1903, vier Jahre nach Marshs Tod, stellte das USNM den Paläontologen Charles W. Gilmore ein, um dessen umfangreiche Sammlung zu bearbeiten. Erst nach zehn Jahren intensiver Forschungsarbeit gelangte Gilmore zu eben jener Kiste, öffnete sie und entdeckte zu seiner freudigen Überraschung darin das Skelett einer bis dahin unbekannten Dinosaurierart, gewissermaßen zum zweiten Mal. Er beschrieb das Skelett erstmalig im Jahr 1913.4Gilmore, C. W. (1913). A new dinosaur from the Lance Formation of Wyoming. Smithsonian Miscellaneous Collections, 61(5), 1–5. |
Nachdem es schließlich vollständig präpariert worden war, veröffentlichte Gilmore 1915 eine umfassende Beschreibung des Exemplars, das seither unter der Inventarnummer USNM 7757 geführt wird. Darin interpretierte er Thescelosaurus als ein agiles, zweibeiniges Tier war, das an schnelles Laufen angepasst gewesen sei. Er fertigte auch ein Lebensbildmodell an, das einen leichten und beweglichen Körper mit schlanken Hinterbeinen zeigte.

 Barnum Brown Barnum Brown |
Gilmore ordnete der Art sechs weitere Fossilien zu, darunter ein Schulterblatt mit Rabenbein, einen Halswirbel und einen Zehenknochen sowie drei Teilskelette, die von Barnum Brown entdeckt worden waren und sich im American Museum of Natural History (AMNH) befanden. |
Hals und Schädel waren jedoch weiterhin unbekannt. Für seine Skelett- und Lebensrekonstruktionen ergänzte Gilmore diese fehlenden Körperteile auf Grundlage des1849 in England gefundenen Hypsilophodon, den er als nahen Verwandten betrachtete. Der neu gewählte Gattungsname leitet sich von den griechischen Wörtern θέσκελος (theskelos = „wunderbar“, „bewundernswert“) und σαῦρος (sauros = „Echse“ oder „Reptil“) ab. Der Artname stammt aus dem Lateinischen und bedeutet „vernachlässigt“, weil der wunderbare Fund so viele Jahre unbeachtet geblieben war.5Gilmore, C. W. (1915). Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming. Proceedings of the United States National Museum, 49(2127), 591–616. doi:10.5479/si.00963801.49-2127.591
Ausstellung
Für die Ausstellung des Skeletts im Museum beließ Gilmore das Skelett weitgehend in seiner ursprünglichen Lage und Unvollständigkeit. Lediglich das rechte Bein, das leicht verrutscht war, wurde neu positioniert. Kleinere Beschädigungen an den Knochen wurden ergänzt, jedoch heller bemalt als die Originalknochen, sodass sich echte und rekonstruierte Bereiche optisch unterscheiden ließen. 1963 wurde das ausgestellte Skelett in eine Wandmontage integriert, zusammen mit den großen Entenschnabel-Dinosauriern Edmontosaurus und Corythosaurus sowie dem Theropoden Gorgosaurus. 1981 wurde die Ausstellung wieder umgestaltet, wobei Thescelosaurus nun höher und weniger gut sichtbar positioniert wurde. Im Zuge von Renovierungsarbeiten zwischen 2014 und 2019 wurden Thescelosaurus und andere ausgestellte Dinosaurier entfernt und durch Gipsabgüsse ersetzt, damit die Originalfossilien weiter präpariert und wissenschaftlich untersucht werden konnten.


Weitere Exemplare und Arten
 William Parks William Parks |
1926 beschrieb William Parks eine neue Art namens Thescelosaurus warreni auf Grundlage eines gut erhaltenen Skeletts aus Alberta in Kanada.6Parks, W. A. (1926). Thescelosaurus warreni, a new species of orthopodous dinosaur from the Edmonton Formation of Alberta. University of Toronto Studies, Geological Series, 21, 1–42. Dieses Skelett unterschied sich aber so deutlich von T. neglectus, weshalb Charles M. Sternberg es 1937 in eine neue Gattung Parksosaurus stellte, die er nach Parks benannte.7Sternberg, C. M. (1937). Classification of Thescelosaurus, with a description of a new species. Geological Society of America, Proceedings of the Paleontological Society, 1936, 375. |

 Charles M. Sternberg Charles M. Sternberg |
1940 benannte Charles M. Sternberg eine weitere Art, Thescelosaurus edmontonensis, basierend auf einem zusätzlichen artikulierten Skelett, welches er selbst in der Edmonton Gruppe bei Rumsey in Alberta entdeckt hatte. Dieses trägt noch immer die Inventarnummer NMC 8537. In seiner Beschreibung äußerte er Zweifel an dem agilen Körperbau, den Gilmore vorgeschlagen hatte. Sternberg argumentierte, dass das Skelett und insbesondere die Gliedmaßen sehr robust gebaut gewesen seien. Sein eigenes Modell von T. edmontonensis zeigte deshalb deutlich kräftigere und muskulösere Beine. 8Sternberg, C. M. (1940). Thescelosaurus edmontonensis, n. sp., and Classification of the Hypsilophodontidae. Journal of Paleontology, 14(5), 481–494. Sternberg hatte dieses Exemplar bereits 1926 erwähnt, doch damals war es noch nicht präpariert.9Sternberg, C. M. (1926). Notes on the Edmonton Formation of Alberta. The Canadian Field-Naturalist, 40(5), 102–105. doi:10.5962/p.338660 |

Das Exemplar umfasst den Großteil der Wirbelsäule, das Becken, die Beine, Schulterblatt und Coracoid, den Arm sowie auch mehrere Knochen des Schädeldachs und einen vollständigen Unterkiefer, der erste überhaupt bekannte von Thescelosaurus. Was zu Sternbergs Zeit noch „Edmonton Formation“ genannt wurde, ist heute allerdings in vier separate Formationen aufgeteilt. Dabei stammt Parksosaurus warreni noch aus der älteren Horseshoe Canyon Formation, während Thescelosaurus edmontonensis aus der jüngeren Scollard Formation bekannt ist.10Eberth, D. A. & Kamo, S. L. (2019). High-precision U-Pb CA-ID-TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River valley, Alberta, Canada. Canadian Journal of Earth Sciences, 57(10), 1220–1237. doi:10.1139/cjes-2019-0019
1974 überarbeitete Peter M. Galton die Gattung Thescelosaurus und beschrieb zusätzliche Fossilien, wodurch die Zahl der bekannten Exemplare auf insgesamt 15 anstieg. Dazu gehören nun auch vier Exemplare aus der Hell Creek Formation, die Barnum Brown schon 1906 und 1909 in Montana entdeckt hatte und von denen einige bereits 1915 von Gilmore erwähnt worden waren.
 Peter M. Galton Peter M. Galton |
1974 überarbeitete Peter M. Galton die Gattung Thescelosaurus und beschrieb zusätzliche Fossilien, wodurch die Zahl der bekannten Exemplare auf insgesamt 15 anstieg. Dazu gehören nun auch vier Exemplare aus der Hell Creek Formation, die Barnum Brown schon 1906 und 1909 in Montana entdeckt hatte und von denen einige bereits 1915 von Gilmore erwähnt worden waren. |
Ein weiteres Exemplar hatten Jacob L. Wortman und Olof A. Peterson an einem unsicheren Fundort entdeckt, zwei Exemplare stammten aus der Frenchman Formation bei Rocky Creek in Saskatchewan, die 1921 von Levi Sternberg gefunden wurden, sowie zwei isolierte Knochen, ebenfalls aus Saskatchewan. Eines von Browns Exemplaren war nur etwa 1,5 Meter unterhalb der Fort Union Formation gefunden – an der stratigraphisch jüngsten Fundstelle, aus der Dinosaurier bekannt sind. Galton kam zu dem Schluss, dass T. edmontonensis lediglich ein robuster gebautes Individuum von T. neglectus war, möglicherweise sogar das andere Geschlecht des Typusexemplars.11Galton, P. M. (1974). Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification. Journal of Paleontology, 48(5), 1048–1067.
William J. Morris beschrieb 1976 drei weitere Teilskelette. Zwei davon wurden vom Präparator Harli Garbani in der Hell Creek Formation im Garfield County, Montana, entdeckt; das dritte stammt aus einem unbekannten Fundort im Harding County, South Dakota. Das erste Exemplar umfasst Teile der Wirbelsäule und des Beckens sowie Schädelknochen, die zuvor bei Thescelosaurus noch unbekannt waren, darunter die Jochbeine und Teile des Hirnschädels. Das zweite zeigt wiederum Hals- und Rückenwirbel sowie einen nahezu vollständigen Unterschenkel mit einem teilweisen Oberschenkelknochen. Morris kam zu dem Schluss, dass dessen Sprunggelenksanatomie und größere Körpergröße einzigartig seien, und benannte deshalb die neue Art Thescelosaurus garbanii zu Ehren des Entdeckers Garbani.12Morris, W. J. (1976). Hypsilophodont dinosaurs. A new species and comments on their systematics. In Churcher, C. S. (Hrsg.), Athlon. Essays on Palaeontology in honour of Loris Shano Russell. Royal Ontario Museum, Life Sciences Miscellaneous Publications, 93–113.13Sues, H.-D. (1980). Anatomy and relationships of a new hypsilophodontid dinosaur from the Lower Cretaceous of North America. Palaeontographica Abteilung A, 169(1–3), 51–72.
Morris argumentierte außerdem, dass das Sprunggelenk von T. edmontonensis, das Galton zuvor als beschädigt und fehlinterpretiert angesehen hatte, tatsächlich deutlich von T. neglectus abweiche und eher Ähnlichkeiten mit T. garbanii zeige. Daher schlug er vor, dass T. edmontonensis und T. garbanii möglicherweise auch einer neuen Gattung außerhalb von Thescelosaurus zugeordnet werden könnten. Das dritte Exemplar umfasst einen großen Teil des Schädels, einige teilweise erhaltene Rückenwirbel und zwei Fingerknochen. Diese Bereiche überschneiden sich jedoch nicht mit den diagnostischen Regionen des Holotypus von T. neglectus, wodurch direkte Vergleiche erschwert werden. Morris ordnete das Exemplar daher nur vorläufig Thescelosaurus zu, vermutete jedoch, dass es eine neue Art repräsentieren könnte. Dieses potenzielle Taxon wurde später als „Hell Creek-Hypsilophodontid“ bezeichnet.
Revisionen und der Weg zur heute gültigen Systematik
Peter M. Galton überarbeitete Thescelosaurus 1995 ein zweites Mal. Er argumentierte, dass die angeblich diagnostischen Merkmale des Sprunggelenks des Exemplars von T. edmontonensis auf Beschädigungen zurückzuführen seien. Dies werde durch das zuvor unbeschriebene linke Sprunggelenk desselben Exemplars belegt, das dieselbe Anatomie wie T. neglectus zeigt. Folglich synonymisierte er T. edmontonensis mit T. neglectus. Galton bestätigte außerdem, dass Morris das Sprunggelenk von T. garbanii korrekt interpretiert hatte, und schlug vor, dass diese Art möglicherweise in eine eigene Gattung gestellt werden sollte. Galton kam zudem zu dem Schluss, dass der Schädel von Morris drittem beschriebenen Exemplar deutlich von Thescelosaurus abweicht. Deshalb benannte er das neue Taxon Bugenasaura infernalis. Der Name bedeutet „Höllische Großwangenechse“, wobei sich der Artname auf die unteren Schichten der Hell Creek Formation bezieht, aus denen das Fossil stammt. Galton ordnete außerdem den Holotypus von T. garbanii der neuen Gattung zu. Er betonte jedoch, dass zusätzliches Fossilmaterial notwendig sei, um diese Zuordnung sicher zu bestätigen.14Galton, P. M. (1995). The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 198(3), 297–311. doi:10.1127/njgpa/198/1995/297 Sollte sich die Zuordnung als korrekt erweisen, hätte der Name garbanii vor infernalis jedoch Priorität.
In seiner Revision von 1995 ordnete Galton außerdem isolierte Zähne aus der campanischen Judith River Formation in Montana der verwandten Gattung Orodromeus zu. Diese Zähne waren 1972 von Ashok Sahni als Thescelosaurus cf. neglectus bestimmt worden, was den damals ältesten Nachweis von Thescelosaurus dargestellt hätte.15Sahni, A. (1972). „The vertebrate fauna of the Judith River Formation, Montana“. Bulletin of the American Museum of Natural History. 147 (6): 321–412.

In einer Studie zur Anatomie von Bugenasaura aus dem Jahr 1999 ordnete Galton außerdem einen Zahn aus der Sammlung des University of California Museum of Paleontology (UCMP 49611) dieser Gattung zu. Besonders bemerkenswert ist, dass dieser Zahn angeblich aus der spätjurassischen Kimmeridge Clay Formation bei Weymouth in England stammt und damit rund 70 Millionen Jahre älter wäre als der Holotyp von Bugenasaura, und zudem von einem anderen Kontinent. Galton vermutete daher, dass das Fossil möglicherweise falsch etikettiert worden war und tatsächlich aus der Lance Formation in Wyoming stammt.16Galton, P. M. (1999). Cranial anatomy of the hysilophodontid dinosaur Bugenasaura infernalis (Ornithischia: Ornithopoda) from the Upper Cretaceous of North America. Revue de Paléobiologie, 18(2), 517–534. Allerdings wurde der Zahn bereits gesammelt, bevor das Museum überhaupt in der Lance-Region aktiv war. Insofern stellt dieser Zahn immer noch ein paläontologisches Rätsel dar. Paul M. Barrett und Susannah Maidment klassifizierten ihn deshalb 2011 als „umbestimmter Ornithischier“.17Barrett, P. M. & Maidment, S. C. R. (2011). Dinosaurs of Dorset. Part III, the ornithischian dinosaurs (Dinosauria, Ornithischia) with additional comments on the sauropods. Proceedings of the Dorset Natural History and Archaeological Society, 132, 145–163.
„Willo“ – Der Dino mit einem Herz aus Stein?
Bei einigen neuen Exemplaren waren nun endlich auch vollständige Schädel erhalten. Besonders bemerkenswerte neue Einblicke eröffnete ein nahezu vollständiges Skelett, das Michael Hammer 1999 in der oberen Hell Creek Formation im Harding County von South Dakota entdeckte. Das Exemplar trägt seither die Inventarnummer NCSM 15278, ist aber auch unter dem Spitznamen „Willo“ bekannt. Besonders bemerkenswert an diesem Exemplar sind Strukturen im Brustbereich, die von einigen Experten als Abdrücke eines komplexen, vogelähnlichen Vierkammerherzens gedeutet wurden.18Fisher, P. E., et al. (2000). Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. Science, 288(5465), 503–505. doi:10.1126/science.288.5465.503 Diese Struktur galt daher als wichtiger Beleg dafür, dass Dinosaurier, wenigstens aber kleine Ornithopoden wie Thescelosaurus, einen endothermen und daher gleichwarmen Stoffwechsel betrieben und agile Tiere mit hohem Energiebedarf waren.
Doch diese Interpretation blieb nicht unwidersprochen. Eine 2001 erschienene Studie wies die Hypothese zurück, da das vermeintliche Herz teilweise eine der Rippe umschließe und an einigen Stellen konzentrisch geschichtete innere Strukturen zeigt, was völlig untypisch für Herzen ist. Stattdessen interpretierten sie das Objekt als Eisen-Konkretion. Solche Konkretionen sind in vergleichbaren Sedimenten häufig, und eine weitere, sehr ähnliche Konkretion befindet sich sogar direkt hinter dem rechten Bein des Exemplars.19Rowe, T., McBride, E. F. & Sereno, P. C. (2001). Technical comment. Dinosaur with a heart of stone. Science, 291(5505), 783a. doi:10.1126/science.291.5505.783a Dale A. Russel und Kollegen verteidigten jedoch die ursprüngliche Interpretation und argumentierten, dass diese Konkretion einzigartig sei und sich um das tatsächliche Herz herum gebildet haben könne.20Russell, D. A., Fisher, P. E., Barrick, R. E. & Stoskopf, M. K. (2001). Reply. Dinosaur with a heart of stone. Science, 291(5505), 783a. doi:10.1126/science.291.5505.783a

 Mary H. Schweitzer Mary H. Schweitzer |
2011 untersuchte ein Forscherteam unter Leitung von Mary H. Schweitzer die Struktur mit mehreren modernen Analyseverfahren erneut. Dazu gehörten fortschrittliche CT-Scans, Histologie, Röntgendiffraktion, Röntgenphotoelektronenspektroskopie sowie Rasterelektronenmikroskopie. |
Schweitzers Team stellte fest, dass die innere Struktur des Objekts wohl keine separaten Herzkammern enthält, sondern aus drei voneinander getrennten Bereichen mit geringerer Dichte besteht, die nicht mit dem Herzen eines Straußes vergleichbar seien. Die angeblichen Wände bestehen aus sedimentären Mineralien, die in biologischen Systemen nicht gebildet werden, darunter Goethit, Feldspäte, Quarz und Gips, außerdem waren Pflanzenfragmente vorhanden. Die für lebende Organismen wichtigen chemischen Elemente Kohlenstoff, Stickstoff und Phosphor konnten in den Proben nicht nachgewiesen werden, und typische Herz-Zellstrukturen fehlten ebenfalls. Lediglich ein kleiner Bereich zeigte möglicherweise tierische Zellstrukturen. Die Autoren kamen hierbei zu dem Schluss, dass ihre Daten die Interpretation als geologische Konkretion deutlich besser stützen als die Deutung als Herz, wobei sie die Möglichkeit offenließen, dass isolierte Gewebereste aber durchaus erhalten geblieben sein könnten.21Cleland, T. P., Stoskopf, M. K. & Schweitzer, M. H. (2011). Histological, chemical, and morphological reexamination of the „heart“ of a small Late Cretaceous Thescelosaurus. Naturwissenschaften, 98(3), 203–211. doi:10.1007/s00114-010-0760-1
Weitere taxonomische Studien
 Clint Boyd Clint Boyd |
2009 überprüften Clint Boyd und Kollegen sowohl die altbekannten Exemplare als auch neuentdecktes Material erneut. Willo brachte aufgrund seiner Vollständigkeit neues Licht in die taxonomischen Debatten. Sein Schädel ist außerdem deutlich flacher und länger als zuvor bei Thescelosaurus angenommen. Die Vergleichsanalyse konnte bestätigen, dass Bugenasaura infernalis definitiv eine Art von Thescelosaurus ist. Die Autoren betrachteten sie jedoch als nicht eindeutig diagnostizierbar.22Boyd, C. A., et. al. (2009). Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura. Journal of Vertebrate Paleontology, 29(3), 758–770. doi:10.1671/039.029.0328 |
 Caleb M. Brown Caleb M. Brown |
Zuletzt beschrieben Caleb M. Brown und Kollegen 2011 eine neue Art, nämlich Thescelosaurus assiniboiensis, auf Grundlage eines Exemplars, das Albert Swanston, ein Museumstechniker des Royal Saskatchewan Museum, 1968 in der Frenchman Formation entdeckt hatte. |
Der ursprünglich angegebene Fundort erwies sich später jedoch als falsch. Bei einer erneuten Untersuchung des Frenchman-River-Tals in den 1980er Jahren kam anhand von Knochen- und Gipsresten heraus, dass die Ausgrabung tatsächlich auf der Nordseite des Tals stattgefunden haben musste, etwa auf halber Höhe der freiliegenden Tonsteinschichten. Zum Zeitpunkt seiner Entdeckung lag das Fossil noch artikuliert vor, wobei der Schwanz bereits aus einem Hang herauswitterte. Es handelt sich um ein kleines Exemplar mit einem fragmentarischen Schädel, dem Großteil der Wirbelsäule, den Beckengürteln und den Hinterbeinen. Der Artname assiniboiensis leitet sich vom historischen Distrikt Assiniboia ab, der den südlichen Teil Saskatchewans umfasste, in dem die Lagerstätte aufgeschlossen ist. Der Name des Distrikts wiederum geht auf die Assiniboine-Völker zurück.23Brown, C. M., Boyd, C. A. & Russell, A. P. (2011). A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America. Zoological Journal of the Linnean Society, 163(4), 1157–1198. doi:10.1111/j.1096-3642.2011.00735.x

Heutige Klassifikation
Die Kontroversen bei der Klassifikation der vielen möglichen Thescelosaurus-, Parksosaurus oder Bugenasaura-Arten bestehen vor allem deshalb, weil die gefundenen Exemplare nur dann direkt miteinander verglichen werden können, wenn dieselben Knochen erhalten sind. Häufig fehlt jedoch überlappendes Material, weshalb die Zuordnung vieler Thescelosaurus-Fossilien lange unsicher blieb. Diese Situation verbesserte sich erst 2014, als Boyd und Kollegen ein neues Exemplar aus der Hell Creek Formation im Dewey County, South Dakota beschrieben. Das Fossil war zuvor von Bill Alley auf Privatland entdeckt und anschließend dem Timber Lake and Area Museum gespendet worden. Obwohl das Exemplar noch nicht vollständig präpariert war, umfasst es einen weitgehend vollständigen, wenn auch leicht zerdrückten Schädel sowie große Teile des restlichen Skeletts. Durch diesen Fund konnten sowohl dieses Exemplar als auch das „Willo“-Exemplar sicher T. neglectus zugeordnet werden.24Boyd, C. A. (2014). The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus. PeerJ, 2, e669. doi:10.7717/peerj.669
Neben der Typusart T. neglectus gelten heute die deutlich größere Art Thescelosaurus garbanii und Thescelosaurus assiniboiensis gültig. Die anderen beiden Arten, T. edmontonicus und T. infernalis sind sogenannte Nomina dubia, also zweifelhafte Namen, die sehr wahrscheinlich zu T. garbanii gehören, wie auch die Gattung Bugenasaura. Die einige Millionen Jahre ältere Gattung Parksosaurus hingegen könnte die Schwestergattung von Thescelosaurus sein und möglicherweise auch sein direkter Vorfahr.25Fonseca, A. O. et al. (2024). A comprehensive phylogenetic analysis on early ornithischian evolution. Journal of Systematic Palaeontology, 22(1), 2346577. doi:10.1080/14772019.2024.2346577 In Die Weißen Steine habe ich mich jedoch an der These orientiert, dass der kleine, grazile T. neglectus und der robustere und größere T. garbanii unterschiedliche Geschlechter repräsentieren könnten. In meiner Romanreihe stellt T. neglectus daher das Weibchen und T. garbanii das Männchen einer gemeinsamen Art dar. Da T. assiniboiensis bislang nur von einem einzigen Exemplar aus Kanada bekannt ist, kommt er in der Geschichte nicht vor.

Verbreitung
Die Unterschiede zwischen den Arten sind allerdings sowohl zeitlich als auch geographisch zu erklären:
- T. neglectus ist aus der Lance Formation in Wyoming sowie der Hell Creek Formation in South Dakota bekannt.
- T. garbanii stammt dagegen nur aus der Hell Creek Formation in Montana.
- Der Holotyp von „T. edmontonensis“, der vermutlich auch zu T. garbanii gehört, wurde in der kanadischen Scollard Formation in Alberta gefunden.
- T. assiniboiensis kam wiederum nur in der Frenchman Formation in Saskatchewan vor.
Die Ablagerung der Lance Formation begann vor etwa 69,42 Millionen Jahren; die Scollard Formation und die Frenchman Formation begannen sich ganz am Ende der Kreidezeit, vor etwa 66,88 Millionen Jahren abzulagern. Die Sedimentation der Fundschichten der Hell Creek Formation setzte spätestens vor 67,2 Millionen Jahren ein.26Eberth, D. A. & Kamo, S. L. (2019). High-precision U-Pb CA-ID-TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River valley, Alberta, Canada. Canadian Journal of Earth Sciences, 57(10), 1220–1237. doi:10.1139/cjes-2019-0019 Wenn man danach geht, könnte T. neglectus der älteste Morphytyp sein, der zuerst von T. garbanii und zuletzt von T. assiniboiensis abgelöst wurde.
Weitere Fossilien, die bislang aber nicht sicher einer der Thescelosaurus-Arten zugeordnet werden konnte, stammen aus der Hell Creek Formation in North Dakota, der Laramie Formation in Colorado, den Ferris Formation, Medicine Bow Formation und Almond Formation in Wyoming sowie der Willow Creek Formation in Montana. Alle diese Fundstellen stammen jedoch aus einem ähnlichen Zeitfenster wie die sicher zugeordneten Thescelosaurus-Arten, was eine Zugehörigkeit zu dieser Gattung sehr wahrscheinlich macht. Zweifel besteht jedoch bei der Zugehörigkeit von ebenfalls Thescelosaurus zugeschriebenen Fossilien aus der älteren Horseshoe Canyon Formation in Alberta sowie der Prince Creek Formation in Alaska, die unweit des spätkreidezeitlichen Nordpols abgelagert wurde, allerdings schon während des frühen Maastrichtiums vor etwa 71 bis 69 Ma. Auch wenn es nicht völlig unmöglich ist, dass es sich bei diesen älteren Funden schon um frühe Thescelosaurus-Formen handeln könnte, ist eine Zugehörigkeit zu Parksosaurus aufgrund des höheren Alters doch wahrscheinlicher.27Weishampel, D. B. et al. (2004). Dinosaur Distribution. In Weishampel, D. B., Osmólska, H. & Dodson, P. (Hrsg.), The Dinosauria. University of California Press, 517–606.28Brown, C. M. & Druckenmiller, P. (2011). Basal ornithopod (Dinosauria: Ornithischia) teeth from the Prince Creek Formation (early Maastrichtian) of Alaska. Canadian Journal of Earth Sciences, 48(9), 1342–1354. doi:10.1139/E11-017 Dennoch war Thescelosaurus im westlichen Nordamerika weit verbreitet und kam in allen küstennahen Lebensräumen im nördlichen Teil Laramidias vor.
Auf meiner interaktiven Karte kannst du die verschiedenen Lagerstätten erforschen und mehr über die dort gefundenen Fossilien erfahren!
Verwandtschaft
Wer jedoch die nächsten Verwandten außerhalb der eigenen Gattung waren und zu welcher Familie Thescelosaurus eigentlich gehört, darüber wurde jahrzehntelang gestritten. In seiner Beschreibung von Thescelosaurus aus dem Jahr 1913 betrachtete Gilmore die Gattung als Mitglied der Camptosauridae, zusammen mit Hypsilophodon, Dryosaurus und Laosaurus.29Gilmore, C. W. (1913). A new dinosaur from the Lance Formation of Wyoming. Smithsonian Miscellaneous Collections, 61(5), 1–5. Doch 1915 stellte er Thescelosaurus stattdessen innerhalb der Hypsilophodontidae auf und ordnete nur noch Hypsilophodon gemeinsam mit ihm dieser Familie zu. Viele spätere Autoren übernahmen diese Einordnung innerhalb der Hypsilophodontidae, sie ist noch heute immer wieder zu lesen.30Gilmore, C. W. (1915). Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming. Proceedings of the United States National Museum, 49(2127), 591–616. doi:10.5479/si.00963801.49-2127.591
 Franz von Nopcsa Franz von Nopcsa
|
Doch gab es um die Zugehörigkeit des kleinen Dinosauriers ein langes Hin und Her, das sogar heute noch nicht abschließend geklärt ist. Franz von Nopcsa31Nopcsa, F. (1928). The genera of reptiles. Palaeobiologica, 1, 163–188. und Friedrich von Huene32Huene, F. F. v. (1956). Paläontologie und Phylogenie der Niederen Tetrapoden. Gustav Fischer Verlag, 536. betrachteten Thescelosaurus dagegen 1928 beziehungsweise 1956 weiterhin als nahen Verwandten von Camptosaurus. |

{kind=link}
1937 stellte Charles M. Sternberg Thescelosaurus und die verwandte Gattung Parksosaurus in eine eigene Familie, die Thescelosauridae.33Sternberg, C. M. (1937). Classification of Thescelosaurus, with a description of a new species. Geological Society of America, Proceedings of the Paleontological Society, 1936, 375. Doch 1940 änderte er seine Meinung und ordnete beide Gattungen wieder in der Hypsilophodontidae ein, jedoch als Mitglieder einer eigenen Unterfamilie, der Thescelosaurinae.34Sternberg, C. M. (1940). Thescelosaurus edmontonensis, n. sp., and Classification of the Hypsilophodontidae. Journal of Paleontology, 14(5), 481–494. 1940 umfasste die Hypsilophodontidae noch erst vier Gattungen: den englischen Hypsilophodon, die nordamerikanischen Thescelosaurus und Parksosaurus sowie den afrikanischen Dysalotosaurus. 1966 ordnete Alfred Sherwood Romer den Großteil der kleinen Ornithopoden dieser Familie zu, was sich bis zum Ende des 20. Jahrhundert als Mehrheitsmeinung hielt.35Romer, A. S. (1966). Vertebrate Paleontology. The University of Chicago Press, 370. Dieses Konzept der Hypsilophodontidae als große monophyletische, also natürliche Verwandtschaftsgruppe wurde auch durch frühe kladistische Studien unterstützt, die Thescelosaurus, paradoxerweise trotz seines geologisch jüngsten Alters, als den ursprünglichsten Vertreter der Hypsilophodontidae interpretierten.36Sereno, P. C. (1986). Phylogeny of the Bird-Hipped Dinosaurs (Ornithischia). National Geographic Research, 2(2), 234–256. 1990 umfasste die Hypsilophodontidae in der ersten Ausgabe des umfangreichen und bis heute noch bedeutenden Buches The Dinosauria bereits 13 Gattungen.37Sues, H.-D. & Norman, D. B. (1990). Hypsilophodontidae, Tenontosaurus, Dryosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hrsg.), The Dinosauria. University of California Press, Berkeley, 498–509.
Doch teilten nicht alle Experten diese Auffassung und das Konzept der Hypsilophodontidae als monophyletische Gruppe geriet zunehmend in Kritik. Schon Anatoly K. Rozhdestvensky38Rozhdestvensky, A. K. (1964). Подотряд Ornithopoda, Птиценогие. In Orlov, Y. A. (Hrsg.), Основы палеонтологии [Fundamentals of Paleontology]. НАУКА, 553–572. und Richard A. Thulborn39Thulborn, R. A. (1974). A new heterodontosaurid dinosaur (Reptilia: Ornithischia) from the Upper Triassic Red Beds of Lesotho. Zoological Journal of the Linnean Society, 55(2), 151–175. doi:10.1111/j.1096-3642.1974.tb01591.x hielten die Thescelosauridae erneut für eine eigenständige Familie. Peter M. Galton ordnete Thescelosaurus 1974 anhand der Proportionen der Hinterbeine wiederum den Iguanodontidae zu.40Galton, P. M. (1974). Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification. Journal of Paleontology, 48(5), 1048–1067. Diese Familie erwies sich später jedoch als polyphyletisch, also keine natürliche Verwandtschaftsgruppe. Daher kehrte Galton 1995 wieder zu einer Einordnung innerhalb der Hypsilophodontidae zurück.41Galton, P. M. (1995). The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 198(3), 297–311. doi:10.1127/njgpa/198/1995/297 Rodney Sheetz schlug 1999 vor, dass Hypsilophodontiden lediglich primitive Formen der Ornithopoden seien, also frühe Vertreter der größeren Gruppe, der sie traditionell zugerechnet wurden. In seiner Analyse standen Thescelosaurus, Parksosaurus und Bugenasaura stufenweise näher zu Hypsilophodon und späteren Ornithopoden, bildeten jedoch keine eigene natürliche Gruppe.42Scheetz, R. D. (1999). Osteology of Orodromeus makelai and the phylogeny of basal ornithopod dinosaurs. Dissertation, Montana State University, 1–189.
Andere Studien kamen zu ähnlichen Ergebnissen. Dabei wurden Thescelosaurus bzw. damals noch Bugenasaura als frühe Ornithopoden nahe dem Ursprung der Gruppe interpretiert, manchmal gemeinsam mit Parksosaurus in einer eigenen Klade. Ein Problem bei T. neglectus bestand vor der Revision von Boyd und Kollegen im Jahr 2009 darin, dass viele zugewiesene Exemplare zu unsicher waren, darunter die Abgrenzung von Bugenasaura und die ungeklärte Frage, ob T. edmontonensis tatsächlich eine eigene Art war. Erst nach der taxonomischen Überarbeitung durch Boyd und Kollegen wurden die systematischen Beziehungen von Thescelosaurus und den sogenannten Hypsilophodontiden klarer. Die Autoren fanden Unterstützung für eine größere Gruppe früher Ornithopoden, bestehend aus Thescelosaurus, Parksosaurus, Zephyrosaurus, Orodromeus und Oryctodromeus.43Boyd, C. A., Brown, C. M., Scheetz, R. D. & Clarke, J. A. (2009). Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura. Journal of Vertebrate Paleontology, 29(3), 758–770. doi:10.1671/039.029.0328 Brown und Kollegen kamen 2011 bei der Beschreibung von T. assiniboiensis zu ähnlichen Ergebnissen. Dieselben Autoren bestätigten diese Resultate 2013 erneut und führten daraufhin den Namen Thescelosauridae für die gesamte Gruppe wieder ein. Diese wurde in die überarbeitete Unterfamilie Thescelosaurinae und die neue Unterfamilie Orodrominae gegliedert.44Brown, C. M., Evans, D. C., Ryan, M. J. & Russell, A. P. (2013). New data on the diversity and abundance of small-bodied ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta. Journal of Vertebrate Paleontology, 33(3), 495–520. doi:10.1080/02724634.2013.746229
Andere Studien fanden hingegen keine enge Verwandtschaft zwischen Parksosaurus und Thescelosaurus, sondern interpretierten Parksosaurus als näher mit dem südamerikanischen Gasparinisaura verwandt. Boyd argumentierte jedoch, dass die Anatomie von Parksosaurus falsch interpretiert worden sei und dass Parksosaurus und Thescelosaurus sehr nahe verwandt seien, möglicherweise seien sie sogar direkte Schwestergruppen.45Boyd, C. A. (2014). The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus. PeerJ, 2, e669. doi:10.7717/peerj.669 Die Kladen Thescelosauridae (alternativ auch Parksosauridae genannt) und Thescelosaurinae wurden inzwischen durch zahlreiche phylogenetische Analysen bestätigt, wenn auch nicht von allen Studien. Allerdings besteht weiterhin Uneinigkeit darüber, ob Thescelosaurus und die Thescelosauriden überhaupt zu den Ornithopoden gehören oder noch basalere Neornithischier darstellen. Boyd betonte 2015, dass viele phylogenetische Analysen, die Thescelosaurus einschlossen, entweder keine Marginocephalia berücksichtigten oder keine klaren Ergebnisse lieferten. Deshalb gebe es keinen eindeutigen Beweis dafür, dass Thescelosaurus tatsächlich ein Ornithopode sei. In seiner eigenen Analyse standen Thescelosaurus und die Thescelosauridae außerhalb der Ornithopoda und bildeten stattdessen eine größere Gruppe nicht-ornithopoder Neornithischier. Boyd kam dabei zu dem Schluss, dass sich Orodrominae und Thescelosaurinae bereits spätestens im Aptium Nordamerikas voneinander getrennt hatten, wobei sich die Orodrominae innerhalb Nordamerikas diversifizierten. Die Thescelosaurinae könnten sich entweder in Nordamerika oder in Asien entwickelt haben.46Boyd, C. A. (2015). The systematic relationships and biogeographic history of ornithischian dinosaurs. PeerJ, 3, e1523. doi:10.7717/peerj.1523
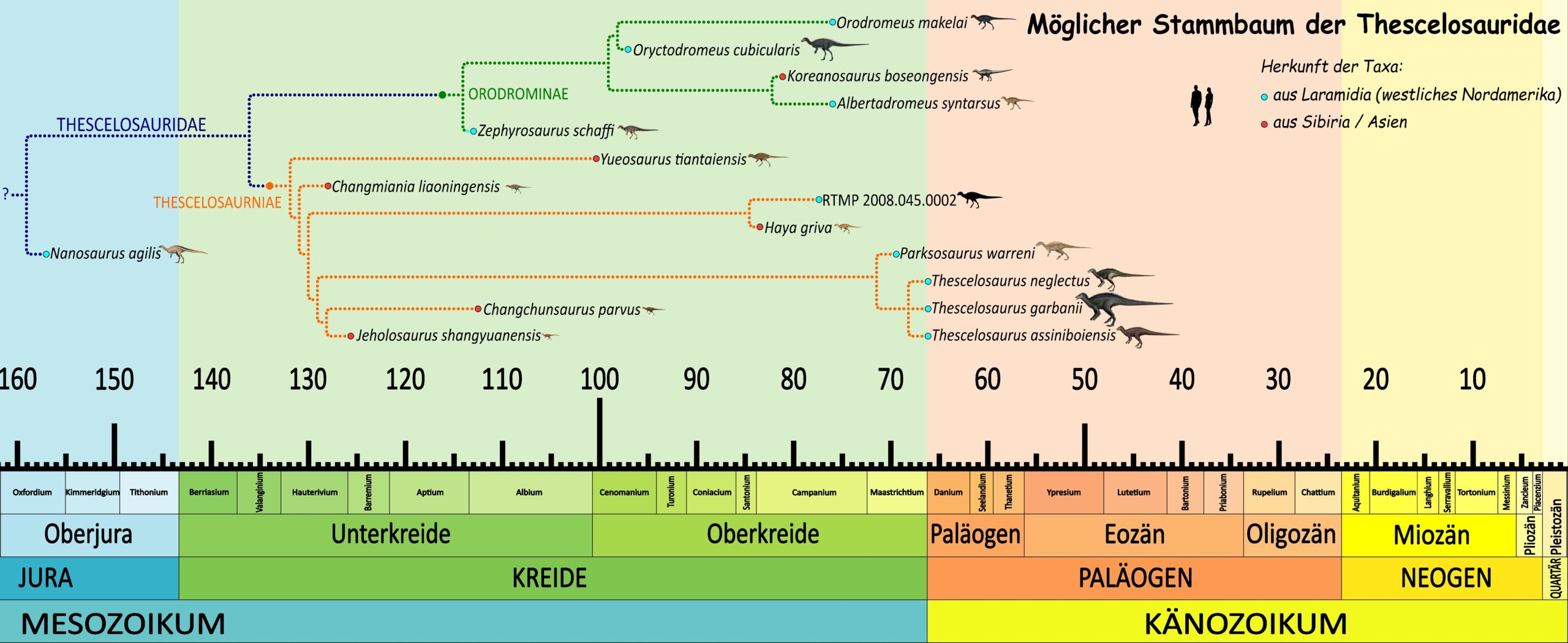
Der Stammbaum der pflanzenfressenden, ungepanzerten Vogelbeckendinosaurier ist seither immer wieder von“ Umbaumaßnahmen“ betroffen. Einige Studien unterstützen diese Position der Thescelosauriden außerhalb der Ornithopoda, andere sehen Thescelosaurus weiterhin innerhalb der Ornithopoden, während manche Analysen überhaupt keine eindeutige Lösung liefern. Die ältesten bekannten Thescelosauriden könnten Changchunsaurus und Zephyrosaurus gewesen sein. Sie stammen allerdings erst aus der mittleren Kreidezeit und wären damit etwa 40 Millionen Jahre jünger als der vermutete Ursprung der Gruppe. Dies deutet auf eine lange sogenannte Geisterlinie hin, also einen geologischen Zeitraum, in dem die Gruppe definitiv existiert haben muss, aber überhaupt keine Fossilien hinterlassen hat.
2024 identifizierten André Fonseca und Kollegen jedoch den spätjurassischen Nanosaurus als den frühesten bekannten Thescelosauriden, wodurch diese Geisterlinie deutlich verkürzt würde. Sie führten in ihrer Studie den Namen Pyrodontia für die Klade ein, die Thescelosaurus mit weiter entwickelten Ornithischiern vereint, wenn die Thescelosauridae außerhalb der Ornithopoda stehen. Der Name bezieht sich auf die frühe und rasche Diversifizierung von Thescelosauriden, Marginocephalia und Ornithopoda.47Fonseca, A.O.; Reid, I.J.; Venner, A.; Duncan, R.J.; Garcia, M.S.; Müller, R.T. (2024). „A comprehensive phylogenetic analysis on early ornithischian evolution“. Journal of Systematic Palaeontology. 22 (1) 2346577. Bibcode:2024JSPal..2246577F. doi:10.1080/14772019.2024.2346577. Die ebenfalls 2024 beschriebene Gattung Fona deutet darauf hin, dass die Thescelosaurinae bereits zu Beginn der späten Kreidezeit fest in Nordamerika etabliert waren.48Avrahami, H. M., Makovicky, P. J., Tucker, R. T. & Zanno, L. E. (2024). A new semi-fossorial thescelosaurine dinosaur from the Cenomanian-age Mussentuchit Member of the Cedar Mountain Formation, Utah. The Anatomical Record, 307(12), 3717–3781. doi:10.1002/ar.25505
Häufigkeit
Thescelosaurus galt lange Zeit als vergleichsweise selten in seinen Paläoökosystemen. Eine Studie aus dem Jahr 1987 schätzte, dass Hypsilophodontiden (einschließlich Thescelosaurus) zusammen mit Pachycephalosauriern nur etwa 2 % der Dinosaurierfaunen der Lance Formation, Hell Creek Formation und Frenchman Formation ausmachten.49Lehman, T. M. (1987). Late Maastrichtian paleoenvironments and dinosaur biogeography in the western interior of North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 60, 189–217. doi:10.1016/0031-0182(87)90032-0 Zwei weitere Zensus-Studien aus den Jahren 199850White, P. D., Fastovsky, D. E. & Sheehan, P. M. (1998). Taphonomy and suggested structure of the dinosaurian assemblage of the Hell Creek Formation (Maastrichtian), eastern Montana and western North Dakota. PALAIOS, 13(1), 41. doi:10.2307/3515280 und 201151Lyson, T. R. & Longrich, N. R. (2011). Spatial niche partitioning in dinosaurs from the latest Cretaceous (Maastrichtian) of North America. Proceedings of the Royal Society B, 278(1709), 1158–1164. doi:10.1098/rspb.2010.1444 kamen zu nur leicht höheren Werten, dass Thescelosaurus etwa 3 % beziehungsweise 5 % der gesamten Dinosaurierfauna der Hell Creek Formation stellte.
Diese niedrigen Werte könnten jedoch auf einen Bias zurückzuführen sein, da spektakulärere Dinosaurier wie Triceratops oder insbesondere Tyrannosaurus bevorzugt gesammelt werden. Heute gilt Thescelosaurus tatsächlich eher als einer der häufigeren Dinosaurier seiner Zeit. Eine Zensusstudie von 2011 in einem Bereich der Hell Creek Formation, in dem Fossilien ohne solche Verzerrungen gesammelt wurden, schätzte den Anteil von Thescelosaurus auf etwa 8 % der gesamten Dinosaurierfauna.52Horner, J. R., Goodwin, M. B. & Myhrvold, N. (2011). Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLOS ONE, 6(2), e16574. doi:10.1371/journal.pone.0016574 Brown und Kollegen kamen 2011 sogar zu dem Schluss, dass Thescelosaurus möglicherweise der häufigste Dinosaurier der Frenchman Formation war und dort etwa 31 % aller Funde ausmachte. Insbesondere an der sogenannten Convenience Store-Fundstelle machten Thescelosaurus-Fossilien sogar 42 % aller Tetrapodenfunde aus. Alle dort gefundenen Exemplare sind sehr klein und vermutlich juvenile Tiere.53Brown, C. M., Boyd, C. A. & Russell, A. P. (2011). A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America. Zoological Journal of the Linnean Society, 163(4), 1157–1198. doi:10.1111/j.1096-3642.2011.00735.x An einer weiteren Fundstelle der Hell Creek Formation, die als Tooth Draw Deposit bekannt ist, entfielen immerhin 22,7 % aller Dinosaurierknochen auf Thescelosaurus.54Stein, W. W. (2021). The Paleontology, Geology and Taphonomy of the Tooth Draw Deposit. Hell Creek Formation (Maastrictian), Butte County, South Dakota. The Journal of Paleontological Sciences, 9, 1–108. Die am häufigsten identifizierbaren Fossilien von Thescelosaurus sind die Zehenknochen des Fußes, während vollständig artikulierte Skelette immer noch sehr selten sind.55Horner, J. (2001). Dinosaurs under the Big Sky. Mountain Press Publications, Missoula, Montana, 128.

Ein Opfer des Meteoriteneinschlags – mit möglicher Hauterhaltung?
2022 berichteten verschiedene Nachrichtenmedien außerdem über ein Thescelosaurus-Exemplar aus der Fossillagerstätte Tanis in North Dakota. Diese Fundstelle gilt als möglicher direkter Nachweis der Auswirkungen des Chicxulub-Asteroideneinschlags im Golf von Mexiko, der zum Kreide-Paläogen-Massenaussterben führte. Das Exemplar könnte, wie Rußspuren und verbrannte Körperregionen nahelegen, tatsächlich infolge des Einschlags verendet sein. Hautabdrücke an seinem Bein seien zudem so gut erhalten, dass sie ein feines Schuppenmuster zeigen.56Amos, J. (2022). Tanis. Fossil found of dinosaur killed in asteroid strike, scientists claim. BBC News.57Martin, S. (2022). Fossil of dinosaur killed in asteroid strike discovered in North Dakota, scientists say. phys.org.
Allerdings ist trotzdem noch unklar, ob und welche Körperbedeckung Thescelosaurus besaß. Schon Gilmore beschrieb 1915 karbonisierte Materialreste nahe den Schultern als mögliche Hautabdrücke mit einer punktierten Struktur, jedoch ohne erkennbares regelmäßiges Muster.58Gilmore, C. W. (1915). Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming. Proceedings of the United States National Museum, 49(2127), 591–616. doi:10.5479/si.00963801.49-2127.591Morris vermutete später sogar eine Panzerung in Form kleiner Hautschuppen oder Osteoderme, die seiner Interpretation nach zumindest entlang der Mittellinie des Halses eines Exemplars verliefen.59Morris, W. J. (1976). Hypsilophodont dinosaurs. A new species and comments on their systematics. In Churcher, C. S. (Hrsg.), Athlon. Essays on Palaeontology in honour of Loris Shano Russell. Royal Ontario Museum, Life Sciences Miscellaneous Publications, 93–113. Solche Strukturen wurden jedoch bei anderen artikulierten Thescelosaurus-Exemplaren nie gefunden. Butler und Galton argumentierten deshalb 2008, dass die von Morris beschriebenen Schuppen wahrscheinlich von einem Krokodil stammen, was heute am wahrscheinlichsten gilt.60Butler, R. J. & Galton, P. M. (2008). The „dermal armour“ of the ornithopod dinosaur Hypsilophodon from the Wealden (Early Cretaceous: Barremian) of the Isle of Wight. A reappraisal. Cretaceous Research, 29(4), 636–642. doi:10.1016/j.cretres.2008.02.002 Aufgrund des Vorhandenseins von Filamenten bei anderen kleinen Ornithopoden wie Kulindadromaeus ist es aber auch wahrscheinlich, dass Thescelosaurus ebenfalls eine Körperbedeckung besessen haben könnte.61Godefroit, P. et al. (2014). A Jurassic ornithischian dinosaur from Siberia with both feathers and scales. Science, 345(6195), 451–455. doi:10.1126/science.1253351 Aus diesem Grund ist er in meiner Romanreihe von einem solchen Filamentkleid bedeckt, ansonsten aber schuppig.
Sinnesleistungen
Die sensorischen und kognitiven Fähigkeiten von Thescelosaurus konnten inzwischen anhand eines CT-Scans des Schädels des „Willo“-Exemplars gut untersucht werden. Obwohl das Gehirn selbst nicht erhalten ist, kann der Schädelinnenraum, der sogenannte Endocast, untersucht werden, in dem das Gehirn einst lag. Insgesamt war das Gehirn im Vergleich zu den meisten anderen Neornithischiern relativ klein, ähnelte aber in seiner Größe dem von Ceratopsiern wie Triceratops. Die kognitiven Fähigkeiten könnten daher wahrscheinlich im Bereich heutiger Reptilien gelegen haben, was Thescelosaurus nicht gerade zu einer Intelligenzbestie machte. Die Forscher kamen bei ihrer Analyse auch zu dem Ergebnis, dass Thescelosaurus weniger stark an schnelles Laufen angepasst war als andere Thescelosauriden, jedoch zwei typische Merkmale von zweibeinigen Lauftieren besaß. Das erste war der vierte Trochanter, ein Knochenkamm am Oberschenkelknochen, an dem der wichtigste Fortbewegungsmuskel ansetzte. Dieser Kamm lag relativ weit oben am Knochen, was schnellere Bewegungen auf Kosten der Kraftentwicklung ermöglichte. Das zweite Merkmal betrifft das Innenohr: Einer der drei Bogengänge, der vordere semicirculäre Kanal, war stark vergrößert. Dies deutet auf einen besonders empfindlichen Gleichgewichtssinn hin, was wiederum auf hohe Beweglichkeit schließen lassen könnte.

Die eher mäßigen geistigen Fähigkeiten könnten darauf hindeuten, dass die sozialen Interaktionen vergleichsweise einfach waren, aber dass Thescelosaurus schon ein gewisses Sozialverhalten an den Tag legte und möglicherweise auch in kleineren Gruppen lebte. An Fundstellen des verwandten Oryctodromeus werden meist zwei bis drei Individuen gemeinsam gefunden, was möglicherweise die typische Gruppengröße dieser Tiere widerspiegelt. Thescelosaurus könnte ebenfalls in solchen kleinen Gruppen gelebt haben, wobei die Belege dafür bislang aber nur schwach sind. Immerhin kann eine Kommunikation mit differenzierten Lauten, also eine gewisse Form der „Sprache“, recht sicher ausgeschlossen werden: Das Hörvermögen von Thescelosaurus war vermutlich eher schlecht. Der optimale Hörbereich wird auf etwa 296 bis 2150 Hertz geschätzt und war damit deutlich enger als bei verwandten Gattungen wie Dysalotosaurus. Der Geruchssinn hingegen war offenbar sehr gut entwickelt. Darauf weisen die großen Riechkolben des Gehirns hin, die etwa 3 % des gesamten Endocast-Volumens ausmachten. Dies entspricht eher heutigen Nagetieren und Hasenartigen und übertrifft den Geruchssinn moderner Vögel.62Button, D. J. & Zanno, L. E. (2023). Neuroanatomy of the late Cretaceous Thescelosaurus neglectus (Neornithischia: Thescelosauridae) reveals novel ecological specialisations within Dinosauria. Scientific Reports, 13(1), 19224. doi:10.1038/s41598-023-45658-3

Ein Tunnelgräber?
Die Kombination dieser Körper- und Sinnesmerkmale lässt eine interessante Interpretation zu. Denn möglicherweise hängen der spezielle Bau des Innenohrs und der Gleichgewichtssinn, sowie auch die Kombination aus schlechtem Hörvermögen und ausgeprägtem Geruchssinn mit einer tunnelgrabenden Lebensweise zusammen. Die Bogengänge des Innenohrs erlauben außerdem Rückschlüsse auf die gewöhnliche Kopfhaltung. Bei heutigen Tieren liegt der laterale semicirculäre Kanal typischerweise horizontal, wenn sich der Kopf in einer „aufmerksamen“ Haltung befindet. Button und Zanno argumentierten, dass der Kopf von Thescelosaurus leicht aber nach oben geneigt gewesen wäre, wenn dieser Kanal horizontal ausgerichtet war. Dies ähnelt der Kopfhaltung von Dysalotosaurus, unterscheidet sich jedoch von den nach unten geneigten Kopfhaltungen, die für viele andere Ornithischier wie Ceratopsier, Ankylosaurier und Hadrosaurier vorgeschlagen wurden.
Das Tier könnte mit Hilfe seines guten Geruchssinns unterirdische Nahrung wie Wurzeln oder Knollen aufgespürt haben. Auch einige anatomische Merkmale des Skeletts könnten mit Grabeverhalten zusammenhängen, darunter die robusten Vorderbeine und die an ihren Spitzen miteinander verschmolzenen Prämaxillaria, die die Schnauzenspitze verstärkten und das Graben erleichtert haben könnten. Zudem war das Schulterblatt breit, was möglicherweise eine größere Ansatzfläche für Muskeln bot, die beim Grabverhalten wichtig waren. Die relativ große Körpergröße von Thescelosaurus schließt grabendes Verhalten nicht zwingend aus, da auch mit dem nur etwas kleineren Oryctodromeus Grabgänge in Verbindung gebracht wurden und sogar deutlich größere Säugetiere grabend leben können. Alternativ besteht auch die Möglichkeit auf, dass Thescelosaurus seine Anpassungen an das Graben lediglich von grabenden Vorfahren geerbt hat, selbst jedoch nicht aktiv grub. Diese Interpretation wird dadurch unterstützt, dass einige typische Grab-Anpassungen fehlen, die beim nah verwandten Oryctodromeus vorhanden sind. Grabendes Verhalten könnte generell bei Thescelosauriden und anderen frühen Neornithischiern weit verbreitet gewesen sein.63Button, D. J. & Zanno, L. E. (2023). Neuroanatomy of the late Cretaceous Thescelosaurus neglectus (Neornithischia: Thescelosauridae) reveals novel ecological specialisations within Dinosauria. Scientific Reports, 13(1), 19224. doi:10.1038/s41598-023-45658-3
 |
In Die Weißen Steine habe ich diese Spekulationen interessiert aufgegriffen und entsprechend erweitert. Hier graben nur die deutlich kleineren Weibchen unterirdische Bauten, in denen sie ihre Jungen aufziehen. Da ich die mögliche tunnelgrabende Lebensweise von kleinen Dinosauriern so faszinierend fand, können auch meine jungen Leser einen kleinen Orodromeus, einen Verwandten von Thescelosaurus, in seinen Bau begleiten und erleben, wie er dort wohnt. Er kommt in einer Geschichte in Traumreise für Kinder in die Zeit der Dinosaurier vor. |

Ernährung und Lebensweise
Thescelosaurus besaß unterschiedliche Zahntypen. Vorne im Kiefer saßen sechs kleine, spitze Prämaxillarzähne. Hinten besaß er dagegen bis zu zwanzig blattförmige Backenzähne, die sich zwischen Ober- und Unterkiefer unterschieden. Diese Zahnformel ist ein primitives Merkmal unter den Ornithischiern, das sonst nur bei frühen Formen wie Lesothosaurus und Scutellosaurus vorkommt. Anders als bei vielen anderen basalen Ornithischiern fehlten den Prämaxillarzähnen jedoch die kleinen Zacken an den Schneidekanten. Junge Individuen hatten möglicherweise weniger als sechs Prämaxillarzähne. Die unterschiedlichen Zahntypen sowie die schmale Schnauze deuten darauf hin, dass es sich um einen selektiven Pflanzenfresser handelte, der sich von einem ganz bestimmten Pflanzentyp ernährte. Andere, etwa gleich große Pflanzenfresser in seinem Lebensraum wie z.B. die Pachycephalosaurier, waren dagegen wohl weniger wählerisch.64Hudgins, M. N., Currie, P. J. & Sullivan, C. (2021). Dental assessment of Stegoceras validum (Ornithischia: Pachycephalosauridae) and Thescelosaurus neglectus (Ornithischia: Thescelosauridae). Paleoecological inferences. Cretaceous Research, 120, 105058. doi:10.1016/j.cretres.2021.105058
Wie sich Thescelosaurus bewegte und wie schnell er Laufen konnte, war in der Forschung schon seit 1915 ein strittiges Thema. Viele Studien widersprachen Gilmores Vorstellung eines besonders schnellen Läufers und verwiesen auf das robuste Skelett, den verhältnismäßig langen Oberschenkelknochen und die kurzen Unterschenkelknochen. Galton schlug 1974 sogar vor, dass sich Thescelosaurus zumindest gelegentlich quadruped, also auf allen Vieren, fortbewegt haben könnte, da die Arme relativ lang und die Hände breit waren.65Galton, P. M. (1974). Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification. Journal of Paleontology, 48(5), 1048–1067. Jüngere Studien gehen davon aber nicht mehr länger aus. Phil Senter und Jared Mackey kamen 2023 zu dem Schluss, dass eine vierbeinige Haltung zwar theoretisch möglich gewesen wäre. Die Wirbelsäule des Rückens war immerhin nach unten gebogen, sodass die Hände den Boden erreichen konnten, selbst wenn die Hinterbeine gestreckt waren. Allerdings hätten die Finger in dieser Haltung seitlich statt nach vorne gezeigt und sich daher nicht effektiv zum Vortrieb geeignet. Eine regelmäßige quadrupede Fortbewegung gilt deshalb als unwahrscheinlich.66Senter, P. & Mackey, J. (2023). Forelimb motion and orientation in the ornithischian dinosaurs Styracosaurus and Thescelosaurus, and its implications for locomotion and other behavior. Palaeontologia Electronica. doi:10.26879/1289

Quellenangaben:
- 1Weishampel, D. B. et al. (2004). Dinosaur Distribution. In Weishampel, D. B., Osmólska, H. & Dodson, P. (Hrsg.), The Dinosauria. University of California Press, 517–606. ↩︎
- 2Brown, C. M. & Druckenmiller, P. (2011). Basal ornithopod (Dinosauria: Ornithischia) teeth from the Prince Creek Formation (early Maastrichtian) of Alaska. Canadian Journal of Earth Sciences, 48(9), 1342–1354. doi:10.1139/E11-017 ↩︎
- 3Boyd, C. A. (2016). The „overlooked marvelous reptile“. Geo News, 43(1), 2–6. ↩︎
- 4Gilmore, C. W. (1913). A new dinosaur from the Lance Formation of Wyoming. Smithsonian Miscellaneous Collections, 61(5), 1–5. ↩︎
- 5Gilmore, C. W. (1915). Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming. Proceedings of the United States National Museum, 49(2127), 591–616. doi:10.5479/si.00963801.49-2127.591 ↩︎
- 6Parks, W. A. (1926). Thescelosaurus warreni, a new species of orthopodous dinosaur from the Edmonton Formation of Alberta. University of Toronto Studies, Geological Series, 21, 1–42. ↩︎
- 7Sternberg, C. M. (1937). Classification of Thescelosaurus, with a description of a new species. Geological Society of America, Proceedings of the Paleontological Society, 1936, 375. ↩︎
- 8Sternberg, C. M. (1940). Thescelosaurus edmontonensis, n. sp., and Classification of the Hypsilophodontidae. Journal of Paleontology, 14(5), 481–494. ↩︎
- 9Sternberg, C. M. (1926). Notes on the Edmonton Formation of Alberta. The Canadian Field-Naturalist, 40(5), 102–105. doi:10.5962/p.338660 ↩︎
- 10Eberth, D. A. & Kamo, S. L. (2019). High-precision U-Pb CA-ID-TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River valley, Alberta, Canada. Canadian Journal of Earth Sciences, 57(10), 1220–1237. doi:10.1139/cjes-2019-0019 ↩︎
- 11Galton, P. M. (1974). Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification. Journal of Paleontology, 48(5), 1048–1067. ↩︎
- 12Morris, W. J. (1976). Hypsilophodont dinosaurs. A new species and comments on their systematics. In Churcher, C. S. (Hrsg.), Athlon. Essays on Palaeontology in honour of Loris Shano Russell. Royal Ontario Museum, Life Sciences Miscellaneous Publications, 93–113. ↩︎
- 13Sues, H.-D. (1980). Anatomy and relationships of a new hypsilophodontid dinosaur from the Lower Cretaceous of North America. Palaeontographica Abteilung A, 169(1–3), 51–72. ↩︎
- 14Galton, P. M. (1995). The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 198(3), 297–311. doi:10.1127/njgpa/198/1995/297 ↩︎
- 15Sahni, A. (1972). „The vertebrate fauna of the Judith River Formation, Montana“. Bulletin of the American Museum of Natural History. 147 (6): 321–412. ↩︎
- 16Galton, P. M. (1999). Cranial anatomy of the hysilophodontid dinosaur Bugenasaura infernalis (Ornithischia: Ornithopoda) from the Upper Cretaceous of North America. Revue de Paléobiologie, 18(2), 517–534. ↩︎
- 17Barrett, P. M. & Maidment, S. C. R. (2011). Dinosaurs of Dorset. Part III, the ornithischian dinosaurs (Dinosauria, Ornithischia) with additional comments on the sauropods. Proceedings of the Dorset Natural History and Archaeological Society, 132, 145–163. ↩︎
- 18Fisher, P. E., et al. (2000). Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. Science, 288(5465), 503–505. doi:10.1126/science.288.5465.503 ↩︎
- 19Rowe, T., McBride, E. F. & Sereno, P. C. (2001). Technical comment. Dinosaur with a heart of stone. Science, 291(5505), 783a. doi:10.1126/science.291.5505.783a ↩︎
- 20Russell, D. A., Fisher, P. E., Barrick, R. E. & Stoskopf, M. K. (2001). Reply. Dinosaur with a heart of stone. Science, 291(5505), 783a. doi:10.1126/science.291.5505.783a ↩︎
- 21Cleland, T. P., Stoskopf, M. K. & Schweitzer, M. H. (2011). Histological, chemical, and morphological reexamination of the „heart“ of a small Late Cretaceous Thescelosaurus. Naturwissenschaften, 98(3), 203–211. doi:10.1007/s00114-010-0760-1 ↩︎
- 22Boyd, C. A., et. al. (2009). Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura. Journal of Vertebrate Paleontology, 29(3), 758–770. doi:10.1671/039.029.0328 ↩︎
- 23Brown, C. M., Boyd, C. A. & Russell, A. P. (2011). A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America. Zoological Journal of the Linnean Society, 163(4), 1157–1198. doi:10.1111/j.1096-3642.2011.00735.x ↩︎
- 24Boyd, C. A. (2014). The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus. PeerJ, 2, e669. doi:10.7717/peerj.669 ↩︎
- 25Fonseca, A. O. et al. (2024). A comprehensive phylogenetic analysis on early ornithischian evolution. Journal of Systematic Palaeontology, 22(1), 2346577. doi:10.1080/14772019.2024.2346577 ↩︎
- 26Eberth, D. A. & Kamo, S. L. (2019). High-precision U-Pb CA-ID-TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River valley, Alberta, Canada. Canadian Journal of Earth Sciences, 57(10), 1220–1237. doi:10.1139/cjes-2019-0019 ↩︎
- 27Weishampel, D. B. et al. (2004). Dinosaur Distribution. In Weishampel, D. B., Osmólska, H. & Dodson, P. (Hrsg.), The Dinosauria. University of California Press, 517–606. ↩︎
- 28Brown, C. M. & Druckenmiller, P. (2011). Basal ornithopod (Dinosauria: Ornithischia) teeth from the Prince Creek Formation (early Maastrichtian) of Alaska. Canadian Journal of Earth Sciences, 48(9), 1342–1354. doi:10.1139/E11-017 ↩︎
- 29Gilmore, C. W. (1913). A new dinosaur from the Lance Formation of Wyoming. Smithsonian Miscellaneous Collections, 61(5), 1–5. ↩︎
- 30Gilmore, C. W. (1915). Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming. Proceedings of the United States National Museum, 49(2127), 591–616. doi:10.5479/si.00963801.49-2127.591 ↩︎
- 31Nopcsa, F. (1928). The genera of reptiles. Palaeobiologica, 1, 163–188. ↩︎
- 32Huene, F. F. v. (1956). Paläontologie und Phylogenie der Niederen Tetrapoden. Gustav Fischer Verlag, 536. ↩︎
- 33Sternberg, C. M. (1937). Classification of Thescelosaurus, with a description of a new species. Geological Society of America, Proceedings of the Paleontological Society, 1936, 375. ↩︎
- 34Sternberg, C. M. (1940). Thescelosaurus edmontonensis, n. sp., and Classification of the Hypsilophodontidae. Journal of Paleontology, 14(5), 481–494. ↩︎
- 35Romer, A. S. (1966). Vertebrate Paleontology. The University of Chicago Press, 370. ↩︎
- 36Sereno, P. C. (1986). Phylogeny of the Bird-Hipped Dinosaurs (Ornithischia). National Geographic Research, 2(2), 234–256. ↩︎
- 37Sues, H.-D. & Norman, D. B. (1990). Hypsilophodontidae, Tenontosaurus, Dryosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hrsg.), The Dinosauria. University of California Press, Berkeley, 498–509. ↩︎
- 38Rozhdestvensky, A. K. (1964). Подотряд Ornithopoda, Птиценогие. In Orlov, Y. A. (Hrsg.), Основы палеонтологии [Fundamentals of Paleontology]. НАУКА, 553–572. ↩︎
- 39Thulborn, R. A. (1974). A new heterodontosaurid dinosaur (Reptilia: Ornithischia) from the Upper Triassic Red Beds of Lesotho. Zoological Journal of the Linnean Society, 55(2), 151–175. doi:10.1111/j.1096-3642.1974.tb01591.x ↩︎
- 40Galton, P. M. (1974). Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification. Journal of Paleontology, 48(5), 1048–1067. ↩︎
- 41Galton, P. M. (1995). The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 198(3), 297–311. doi:10.1127/njgpa/198/1995/297 ↩︎
- 42Scheetz, R. D. (1999). Osteology of Orodromeus makelai and the phylogeny of basal ornithopod dinosaurs. Dissertation, Montana State University, 1–189. ↩︎
- 43Boyd, C. A., Brown, C. M., Scheetz, R. D. & Clarke, J. A. (2009). Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura. Journal of Vertebrate Paleontology, 29(3), 758–770. doi:10.1671/039.029.0328 ↩︎
- 44Brown, C. M., Evans, D. C., Ryan, M. J. & Russell, A. P. (2013). New data on the diversity and abundance of small-bodied ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta. Journal of Vertebrate Paleontology, 33(3), 495–520. doi:10.1080/02724634.2013.746229 ↩︎
- 45Boyd, C. A. (2014). The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus. PeerJ, 2, e669. doi:10.7717/peerj.669 ↩︎
- 46Boyd, C. A. (2015). The systematic relationships and biogeographic history of ornithischian dinosaurs. PeerJ, 3, e1523. doi:10.7717/peerj.1523 ↩︎
- 47Fonseca, A.O.; Reid, I.J.; Venner, A.; Duncan, R.J.; Garcia, M.S.; Müller, R.T. (2024). „A comprehensive phylogenetic analysis on early ornithischian evolution“. Journal of Systematic Palaeontology. 22 (1) 2346577. Bibcode:2024JSPal..2246577F. doi:10.1080/14772019.2024.2346577. ↩︎
- 48Avrahami, H. M., Makovicky, P. J., Tucker, R. T. & Zanno, L. E. (2024). A new semi-fossorial thescelosaurine dinosaur from the Cenomanian-age Mussentuchit Member of the Cedar Mountain Formation, Utah. The Anatomical Record, 307(12), 3717–3781. doi:10.1002/ar.25505 ↩︎
- 49Lehman, T. M. (1987). Late Maastrichtian paleoenvironments and dinosaur biogeography in the western interior of North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 60, 189–217. doi:10.1016/0031-0182(87)90032-0 ↩︎
- 50White, P. D., Fastovsky, D. E. & Sheehan, P. M. (1998). Taphonomy and suggested structure of the dinosaurian assemblage of the Hell Creek Formation (Maastrichtian), eastern Montana and western North Dakota. PALAIOS, 13(1), 41. doi:10.2307/3515280 ↩︎
- 51Lyson, T. R. & Longrich, N. R. (2011). Spatial niche partitioning in dinosaurs from the latest Cretaceous (Maastrichtian) of North America. Proceedings of the Royal Society B, 278(1709), 1158–1164. doi:10.1098/rspb.2010.1444 ↩︎
- 52Horner, J. R., Goodwin, M. B. & Myhrvold, N. (2011). Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLOS ONE, 6(2), e16574. doi:10.1371/journal.pone.0016574 ↩︎
- 53Brown, C. M., Boyd, C. A. & Russell, A. P. (2011). A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America. Zoological Journal of the Linnean Society, 163(4), 1157–1198. doi:10.1111/j.1096-3642.2011.00735.x ↩︎
- 54Stein, W. W. (2021). The Paleontology, Geology and Taphonomy of the Tooth Draw Deposit. Hell Creek Formation (Maastrictian), Butte County, South Dakota. The Journal of Paleontological Sciences, 9, 1–108. ↩︎
- 55Horner, J. (2001). Dinosaurs under the Big Sky. Mountain Press Publications, Missoula, Montana, 128. ↩︎
- 56Amos, J. (2022). Tanis. Fossil found of dinosaur killed in asteroid strike, scientists claim. BBC News. ↩︎
- 57Martin, S. (2022). Fossil of dinosaur killed in asteroid strike discovered in North Dakota, scientists say. phys.org. ↩︎
- 58Gilmore, C. W. (1915). Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming. Proceedings of the United States National Museum, 49(2127), 591–616. doi:10.5479/si.00963801.49-2127.591 ↩︎
- 59Morris, W. J. (1976). Hypsilophodont dinosaurs. A new species and comments on their systematics. In Churcher, C. S. (Hrsg.), Athlon. Essays on Palaeontology in honour of Loris Shano Russell. Royal Ontario Museum, Life Sciences Miscellaneous Publications, 93–113. ↩︎
- 60Butler, R. J. & Galton, P. M. (2008). The „dermal armour“ of the ornithopod dinosaur Hypsilophodon from the Wealden (Early Cretaceous: Barremian) of the Isle of Wight. A reappraisal. Cretaceous Research, 29(4), 636–642. doi:10.1016/j.cretres.2008.02.002 ↩︎
- 61Godefroit, P. et al. (2014). A Jurassic ornithischian dinosaur from Siberia with both feathers and scales. Science, 345(6195), 451–455. doi:10.1126/science.1253351 ↩︎
- 62Button, D. J. & Zanno, L. E. (2023). Neuroanatomy of the late Cretaceous Thescelosaurus neglectus (Neornithischia: Thescelosauridae) reveals novel ecological specialisations within Dinosauria. Scientific Reports, 13(1), 19224. doi:10.1038/s41598-023-45658-3 ↩︎
- 63Button, D. J. & Zanno, L. E. (2023). Neuroanatomy of the late Cretaceous Thescelosaurus neglectus (Neornithischia: Thescelosauridae) reveals novel ecological specialisations within Dinosauria. Scientific Reports, 13(1), 19224. doi:10.1038/s41598-023-45658-3 ↩︎
- 64Hudgins, M. N., Currie, P. J. & Sullivan, C. (2021). Dental assessment of Stegoceras validum (Ornithischia: Pachycephalosauridae) and Thescelosaurus neglectus (Ornithischia: Thescelosauridae). Paleoecological inferences. Cretaceous Research, 120, 105058. doi:10.1016/j.cretres.2021.105058 ↩︎
- 65Galton, P. M. (1974). Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification. Journal of Paleontology, 48(5), 1048–1067. ↩︎
- 66Senter, P. & Mackey, J. (2023). Forelimb motion and orientation in the ornithischian dinosaurs Styracosaurus and Thescelosaurus, and its implications for locomotion and other behavior. Palaeontologia Electronica. doi:10.26879/1289 ↩︎
Hat dir dieser Beitrag gefallen?
Für noch mehr Lesestoff schau gerne in die Kategorie Meine Bücher. Alle meine Werke findest du auch in der Sidebar rechts. Dort gibt es auch einen Direktlink zu Amazon, wo du sie gleich bestellen kannst. ➡️
Wie kannst du Die Weißen Steine sonst noch unterstützen?
Klar, in erster Linie natürlich, wenn du dir eines meiner Bücher kaufst. Aber du kannst mir auch auf anderen Wegen eine große Freude machen:
- Teile diesen Beitrag auf deinen Social-Media-Kanälen!
- Schreibe mir eine nette Rezension auf Amazon!
- Kaufe hier auf der Website über Affiliate Links ein!
Ich kriege übrigens auch eine kleine Unterstützung, wenn du etwas völlig anderes kaufst. Es muss nicht unbedingt der hier beworbene Artikel sein! Jeder kleine Centbetrag hilft, diese Seite am Laufen zu erhalten.
 |
Zusammen mit meiner lieben Frau Sahar habe ich auch noch eine Menge mehr zu bieten! Sahar ist nämlich eine großartige Künstlerin und fertigt Gemälde, Lesezeichen, Stofftaschen und viele andere coole Artikel an. Auch mit tollen Urzeit-Motiven! Klick dich einfach mal durch die Beiträge in der Kategorie Produkte & Services. Bestimmt findest du da etwas nach deinem Geschmack. Oder auch ein schönes Geschenk für deine Lieben. |
Einen Grund, anderen eine Freude zu bereiten, findet man schließlich immer!