
Name: Edmontosaurus annectens („Verbundene Echse aus Edmonton“)
Beschrieben: 1892 von Othniel C. Marsh
Ordnung: Ornithischia; Familie: Hadrosauridae
Länge: ♂ 13,4m, ♀ bis 12,7m; Ausnahmeexemplare: über 14,5m
Gewicht: ♂ 6.200kg, ♀ bis 6.600kg; Ausnahmeexemplare: über 13.000kg
Ernährung: herbivor
Beschreibung:
Edmontosaurus ist ein intelligentes, tagaktives und sehr soziales Herdentier. Obwohl er sich meist auf allen Vieren fortbewegt, kann sich dieser Dinosaurier auch auf zwei Beine aufrichten. So kann er Äste und Blätter von hohen Bäumen noch in über sechs Metern Höhe abweiden. In seinem breiten, schnabelartigen Maul trägt ein Edmontosaurus eine Batterie aus mehr als 2.000 ständig nachwachsenden Zähnen, mit denen er seine Nahrung zerkaut. Diese Zähne bilden eine Art Reibeplatte, mit der er seine Nahrung zermahlen kann, weit effizienter als jeder andere Dinosaurier seiner Zeit und braucht deswegen in der Nahrungswahl nicht pingelig zu sein. Blätter, Farne, Nadeln, ganze Zweige, Nüsse, Nadelbaumzapfen trennt er mit seinem breiten, verhornten Schnabel ab und kaut zerkaut sie dann zu einem zähen Brei.

Die Männchen haben eine bunt gefärbte Schnauze und einen strahlend roten Kopfkamm. Sie sind außerdem deutlich bulliger gebaut und werden auch größer als die Weibchen. Im Durchschnitt wird ein ausgewachsenes Edmontosaurus-Männchen etwa 13 m lang und vier Tonnen schwer. Besonders starke und ältere Exemplare können aber auch deutlich größer werden und das doppelte Gewicht erreichen. Ansonsten ähneln sich weibliche und männliche Tiere sehr. Sie haben eine hellbraune, beinahe schon gelbliche Haut, die am Bauchbereich immer heller wird. Ihr Schwanz ist darüber mit breiten, dunklen Streifen verziert, die ein individuelles Muster bilden. Sie ermöglichen Edmontosaurus eine gute Tarnung und verwirrt Jäger bei der Verfolgung, wenn die gesamte Herde sich zur Flucht in Bewegung setzt. Dann fällt es selbst einem scharfäugigen Tyrannosaurus schwer, sich auf nur eines der fliehenden Tiere zu konzentrieren.
Lebensweise:
Eine Edmontosaurus-Herde auszumachen ist nicht schwer: Bevor man sie zu Gesicht bekommt, kann man sie schon längst hören. Zur Kommunikation in der Herde geben die Tiere laute, basstrompetenartige Töne von sich. Jedes Tier hat dabei eine individuelle Stimme, die seine Artgenossen über weite Strecken erkennen können. Die Herde nutzt diese Rufe nicht nur, um in dichten Wäldern oder über große Entfernungen zusammenzubleiben, sondern auch, um sich gegenseitig vor Feinden zu warnen. Besonders Wachen am Rand der Gruppe sind immer aufmerksam und schlagen bei Gefahr sofort Alarm. Wenn Nahrungsmangel herrscht, kommt es vor, dass Herden über weite Strecken wandern, um neue Weidegründe zu finden. Dabei überqueren sie Flüsse, Sümpfe und sogar küstennahe Gebiete. Interessant ist, dass trotz des Größenunterschieds die Herden meist von den Weibchen geführt werden. Die Männchen leben oft auch ganz ohne soziale Bande als Einzelgänger. Einige schließen sich nur während der Paarungszeit einer Herde an, während andere ihr ganzes Leben über in kleinen Männchengruppen bleiben.

Wie alle Entenschnabelsaurier sind die Tiere fürsorgliche Eltern, die ihre Eier in großen Nistkolonien ablegen. Im wiederkehrenden Rhythmus schließen sich die Tiere zu einem großen Herdenverbund von oft mehr als zwei Dutzend Tieren zusammen, die gemeinsam ihre Eier ausbrüten und die Jungen in den ersten Wochen noch im Nest füttern. Die schiere Größe ihrer Zahl schreckt dabei die meisten Fressfeinde ab. Wenn er Junge führt, ist Flucht schließlich keine Option mehr. Edmontosaurus kann sich aber auch ohne „Waffen“ gut zur Wehr setzen. Dazu setzt er seinen massigen Körper, die hufbewehrten Vordergliedmaßen oder rückwärts gewandte Tritte ein, wenn ein Jäger ihm zu nahe kommt. Die älteren, größeren Individuen bilden dabei oft eine Art Schutzwall um Jungtiere und Weibchen. Selbst für einen Tyrannosaurus wird die Jagd auf einen dieser Riesen so zu einer großen Herausforderung.
Tyrannosaurus in Die Weißen Steine:
 |
Band II:Der Kadaver eines Edmontosaurus wird von der Schülergruppe im Kapitel „Wertloser Ballast“ entdeckt, unweit eines Walnussbaumes, dessen Früchte den Schülern die ausgehungerten Bäuche endlich wieder füllen. Später im Kapitel werden auch lebende Tiere gesichtet. Eine Edmontosaurus-Herde lebt auch in der Nähe von Johns Höhlenwohnung beim Tal, die er im Kapitel „Zuflucht“ erstmals sichtet und in „Rettung“ als eher unliebsame Nachbarn wahrnimmt. In „Das Band“ haben John und Razor eine Begegnung mit einem aggressiven Männchen, die sie nur um Haaresbreite überleben. Auch die Gruppe um Leon und Marie begegnet hin und wieder einer anderen Herde dieser Entenschnabelsaurier, wie im Kapitel „Der Bergpass“ beschrieben wird. |
 |
Band III:Eine Edmontosaurus-Herde nähert sich Moritz in„Der gehörnte Koloss“ am See, um dort zu saufen. Eine andere Herde lebt in der Nähe von Johns Tal, wo er es in „Dämonen“ schließlich schafft, ein Jungtier zu erlegen und seine beim Sturm verlorenen Vorräte wieder aufzustocken. In „Weiche Matratzen“ begegnen Heinrich und Leon auf dem Rückweg vom Strand der Edmontosaurus-Herde, die sich ihnen gegenüber zwar misstrauisch, aber nicht aggressiv verhält. In „Assisi“ entdecken Alicia und ihre Freundinnen in weiter Ferne ebenfalls eine Herde. Somit ist Edmontosaurus im dritten Band der prominenteste pfalnzenfressende Dinosaurier. |
 |
Band IV:Alina und ihre Freundinnen sichten eine Edmontosaurus-Herde im Kapitel „Freundinnen“. Einige Edmontosaurus befinden sich unter den Tieren, vor die vor dem Vulkanausbruch in „Der sterbende See“ panisch fliehen. Auch Katharina und Lea werden in „Der Weg zurück ins Leben“ Zeuge einer anderen fliehenden Herde, die direkt vor der Mauer des Tals entlangrast, aufgeschreckt durch den gewaltigen Knall der Vulkanexplosion. Marie und ihre Freunde sichten ein Edmontosaurus-Männchen in „Heinkehr“, als sie mit dem bewusstlosen Max über die Ebene wandern. |
 |
Hier geht’s zurück zur Hauptseite Leben in der Urzeit! |

Wissenschaftliches zu Edmontosaurus:
 |
Edmontosaurus annectens war einer der größten und auch häufigsten Dinosaurier seiner Zeit und weit verbreitet in Laramidia. Fossilien, die ihm klar zugewiesen werden können, stammen aus der Lance– und Hell Creek Formation in den USA sowie der kanadischen Frenchman Formation. Dort machen seine Fossilien bis zu einem Siebtel aller gefundenen Dinosaurier aus, was ihn zum dritthäufigsten Dinosaurier nach Triceratops und Tyrannosaurus macht.1Bakker, R. T. (1986). The Dinosaur Heresies. 438.2Lehman, T. M. (2001). Late Cretaceous dinosaur provinciality. In Tanke, D. & Carpenter, K. (Hgg.), Mesozoic Vertebrate Life. Indiana University Press, Bloomington/Indianapolis, 310–328.3Weishampel, D. B. et al. (2004). Dinosaur distribution. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 517–606.4Russell, D. A. (1989). An Odyssey in Time. Dinosaurs of North America. 175–180. |
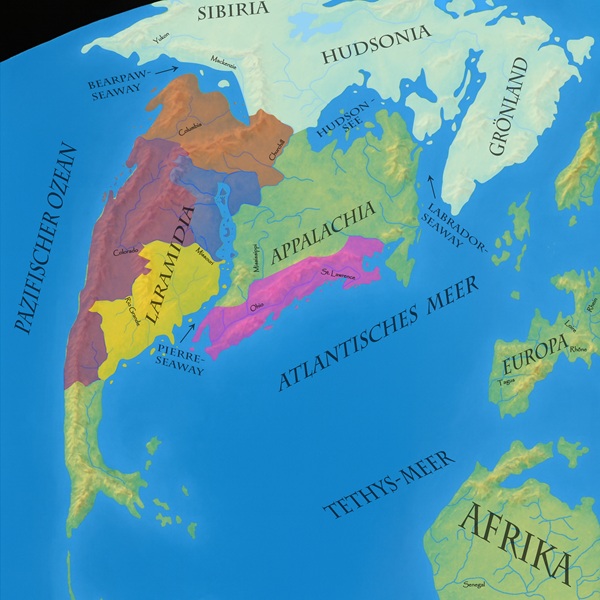
Es ist aber sehr wahrscheinlich, dass er als Art, oder immerhin als Gattung auch sehr viel weiter verbreitet war. Fossilien, die als Edmontosaurus sp. katalogisiert werden, sind aus fast allen Lagerstätten der späten Kreidezeit des westlichen Nordamerika bekannt. Sogar noch im äußersten Norden Alaskas kam die Gattung vor, bis zu den Regionen, wo sich heute der US-Bundesstaat Colorado befindet. Möglicherweise war Edmontosaurus ein wanderndes Tier, das im jährlichen Wechsel von einer Region zur anderen migrierte und dabei vielleicht sogar Asien (Sibiria) erreichte. Er lebte also in den blauen, roten und weißen Regionen auf dieser Karte. Ob er auch weiter westlich und südlich, also in den violetten und gelben Gebieten vorkam, ist derzeit nicht sicher wissenschaftlich belegt.
Forschungsgeschichte
Die Forschungsgeschichte von Edmontosaurus und auch sein taxonomischer Status ist ausgesprochen kompliziert und beginnt eigentlich schon viele Jahrzehnte bevor dieser Gattungsname überhaupt geprägt wurde. Tatsächlich hat kaum ein Dinosaurier seinen „Vornamen“ so häufig gewechselt wie derjenige, den wir in diesem Artikel Edmontosaurus nennen wollen. Denn auch heute noch ist dieser Name alles andere als unumstritten! Hier möchte ich nun Station für Station dieser Geschichte durchgehen und dem Leser vorführen, wie dieses interessante Tier trotzdem zu einem der am besten erforschten Dinosaurier aller Zeiten geworden ist. Tatsächlich wissen wir, selbst wenn wir uns über seinen Namen nicht wirklich einig sind, über kaum einen anderen Bewohner der Hell Creek Formation inzwischen mehr und ausführlicher Bescheid als über ihn! Insofern war er auch das Tier, bei dessen Rekontruktion ich mir in meinem Roman die wenigsten literarischen Freiheiten gelassen habe. Sogar seine Farbe ist uns inzwischen bekannt!
Aber der Reihe nach. Seine Geschichte beginnt wie bei so vielen anderen Bewohnern der Hell Creek Formation auch mit den sogenannten Bone Wars, einer erbitterten Fehde zwischen den zwei führenden Paläontologen des späten 19. Jahrhunderts. Othniel C. Marsh und Edward D. Cope, die sich abgrundtief hassten, versuchten in dieser Zeit sich ständig mit neuen und besonders spektakulären wissenschaftlichen Entdeckungen zu übertrumpfen. Beide beschäftigten Ausgrabungssteams, die im Wilden Westen nach Fossilien suchten.
Diclonius
Das erste bedeutende Exemplar von Edmontosaurus annectens, das heute noch erhalten ist und die Inventarnummer AMNH 5730 trägt, entdeckten Jacob L. Wortman und Robert S. Hill im Jahre 1882, die für Edward D. Cope arbeiteten. Es bestand aus einem vollständigen Schädel und dem größten Teil eines Skeletts und wurde nordöstlich der Black Hills in South Dakota gefunden, was heute ein Gebiet in der Hell Creek Formation ist. Der Fund umfasste ursprünglich auch umfangreiche Abdrücke der Haut. Ein Großteil des Beckens und Teile des Rumpfes fehlten jedoch, da ein Bach das Fossil bereits vor seiner Entdeckung teilweise freigespült und beschädigt hatte. Der Schnabel zeigte Abdrücke einer hornartigen Hülle mit einer zahnähnlichen Reihe ineinandergreifender Strukturen an Ober- und Unterkiefer.5Osborn, H. F. (1909). The Upper Cretaceous iguanodont dinosaurs. Nature, 81(2075), 160–162. doi:10.1038/081160a06Norell, M. A., Gaffney, E. S. & Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. Knopf, New York, 156–158.7Glut, D. F. (1997). Anatotitan. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 132–134.

 Edward D. Cope Edward D. Cope |
Bei seiner Erstbeschreibung ordnete Edward D. Cope das Exemplar der Art Diclonius mirabilis zu. Diesen Namen bildete er, indem er die von ihm zuvor eingeführte Gattung Diclonius mit der älteren Art Trachodon mirabilis kombinierte, die ursprünglich von seinem Lehrer Joseph Leidy auf Grundlage von Zähnen beschrieben worden war. Cope war der Ansicht, dass Leidy die Gattung Trachodon nicht ausreichend charakterisiert hatte und sie später selbst auch gar nicht mehr verwendete. Deshalb überführte er die ältere Art in seine neu geschaffene Gattung Diclonius.8Cope, E. D. (1883). On the characters of the skull in the Hadrosauridae. Proceedings of the Philadelphia Academy of Natural Sciences, 35, 97–107. Nach Copes Tod im Jahr 1897 wurde das Exemplar vom American Museum of Natural History erworben, wo es 1899 auch seine heutige Sammlungsnummer AMNH 5730 erhielt. |
 Joseph Leidy Joseph Leidy |
Joseph Leidy hatte inzwischen auch selbst erkannt, dass seine Gattung Trachodon, die er 1856 beschrieben hatte,9Leidy, J. (1856). Notice of remains of extinct reptiles and fishes, discovered by F. V. Hayden in the Bad Lands of the Judith River, Nebraska Territories. Proceedings of the Academy of Natural Science Philadelphia, 8, 72–73. auf Fossilien mehrerer unterschiedlicher Dinosaurier beruhte. Obwohl er versucht hatte, die Einordnung zu überarbeiten, veröffentlichte er jedoch keine formale taxonomische Neuregelung.10Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.11Marsh, O. C. (1892). Restorations of Claosaurus and Ceratosaurus. American Journal of Science, 44(262), 343–349. doi:10.2475/ajs.s3-44.262.343 |
Trachodon, Hadrosaurus und Claosaurus
 John B. Hatcher John B. Hatcher |
Einer der Knochenjäger, der 1889 im Auftrag von Copes Erzfeind Othniel C. Marsh arbeitete, war John B. Hatcher. Bei einer seiner Expeditionen entdeckte er in der Lance Formation im heutigen Niobrara County in Wyoming, das damals noch zum Converse County gehörte, einen riesigen Unterkiefer eines Dinosauriers. Diesen schickte er zusammen mit vielen anderen Fundstücken zu Marsh an die Yale University. |
 Othniel C. Marsh Othniel C. Marsh |
Marsh benannte dieses Fossil 1890 als Trachodon longiceps. Es trägt heute die Sammlungsnummer YPM 616. Sein langer und schmaler Unterkiefer weist denselben auffälligen seitlichen Knochenkamm auf wie Copes Exemplar. Allerdings ist er deutlich größer. Während der Unterkiefer von Copes Fossil etwa 92 Zentimeter lang ist, wird die Länge von Marshs Exemplar auf rund 110 Zentimeter geschätzt.12Marsh, O. C. (1890). Additional characteristics of the Ceratopsidae, with notice of new Cretaceous dinosaurs. American Journal of Science, 3rd Series, 39(233), 418–426. doi:10.2475/ajs.s3-39.233.418 |
Zwei Jahre später, also 1892, beschrieb Marsh mehrere ebenfalls von Bell gefundene Exemplare als neue Art der Gattung Claosaurus, die er bereits 1890 benannt hatte. Das Typusexemplar USNM 2414 bestand aus einem teilweise erhaltenen Schädeldach und einem Teilskelett. Ein zweites Exemplar mit Schädel und Skelett, YPM 2182, bestimmte Marsh als Paratyp.13Marsh, O. C. (1892). Notice of new reptiles from the Laramie Formation. American Journal of Science, 43(257), 449–453. doi:10.2475/ajs.s3-43.257.449 Dieser „Claosaurus annectens“ ist von besonderer historischer Bedeutung, da er zu den ersten Dinosauriern gehört, für die eine vollständige Skelettrekonstruktion angefertigt wurde. Er war außerdem der erste Hadrosaurier, bei dem dies geschah.14Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.15Marsh, O. C. (1892). Restorations of Claosaurus and Ceratosaurus. American Journal of Science, 44(262), 343–349. doi:10.2475/ajs.s3-44.262.343

Die Exemplare YPM 2182 und USNM 2414 stellen außerdem die ersten beiden nahezu vollständig montierten Dinosaurierskelette in den Vereinigten Staaten dar. YPM 2182 wurde schon 1901 der Öffentlichkeit präsentiert, während USNM 2414 erst im Jahr 1904 ausgestellt wurde.16Lucas, F. A. (1904). The dinosaur Trachodon annectens. Smithsonian Miscellaneous Collections, 45, 317–320.

 Charles H. Sternberg Charles H. Sternberg |
Im Zeitraum von 1902 bis 1915 wurden zwei weitere bedeutende Exemplare von Claosaurus annectens entdeckt. Das erste war das sogenannte „mumifizierte“ Exemplar AMNH 5060, das 1908 von Charles H. Sternberg und seinen Söhnen in Gesteinen der Lance Formation nahe Lusk in Wyoming gefunden wurde. Sternberg arbeitete zu dieser Zeit für das British Museum of Natural History. Henry F. Osborn vom American Museum of Natural History konnte das Fossil jedoch für 2.000 US-Dollar erwerben. |

1910 bargen die Sternbergs in derselben Region ein zweites, ähnliches Exemplar. Dieses war zwar schlechter erhalten, besaß jedoch ebenfalls erhaltene Hautabdrücke. Sie verkauften das Fossil an das Senckenberg Naturmuseum in Frankfurt am Main, wo sich die Mumie heute noch befindet. Sie erhielt dort die Sammlungsnummer SM 4036 und wurde inzwischen auf den Namen „Edmond“ getauft. Nach wie vor gehört dieses Exemplar zu den Herzstücken der Ausstellung im Museum und zu den wichtigsten Forschungsobjekten der Dinosaurierforschung in Deutschland. 17Norell, M. A., Gaffney, E. S. & Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. Knopf, New York, 154–155.18Dal Sasso, C. & Brillante, G. (2004). Dinosaurs of Italy. Indiana University Press, Bloomington and Indianapolis, 112.

 Barnum Brown Barnum Brown |
Ein weiteres nahezu vollständiges Skelett mit der Sammlungsnummer AMNH 5886 wurde 1904 in der Hell Creek Formation am Crooked Creek im zentralen Montana von dem Rancher Oscar Hunter entdeckt. Als Hunter den teilweise freiliegenden Fund machte, stritt er sich mit einem Begleiter darüber, ob die Knochen von einem kürzlich verendeten Tier oder doch fossilen Ursprungs seien. Um seine Meinung zu beweisen, trat Hunter mehrere Wirbelstücke ab und zeigte so ihre steinerne Beschaffenheit, beschädigte dabei jedoch den Fund stark. Ein Cowboy namens Alfred Sensiba tauschte bei Hunter das Fossil gegen eine Pistole ein und verkaufte es anschließend an den berühmten Knochenjäger Barnum Brown. Dieser bedauerte die Beschädigungen sehr, konnte das restliche Skelett aber 1906 für das American Museum of Natural History bergen.19Norell, M. A., Gaffney, E. S. & Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. Knopf, New York, 156–158. |
Dieses Exemplar besaß eine nahezu vollständige Wirbelsäule und ermöglichte dadurch eine wesentlich genauere Rekonstruktion von Copes und Marshs früheren Funden. 1908 wurden beide Skelette nebeneinander im American Museum of Natural History unter dem Namen Trachodon mirabilis ausgestellt. Das Exemplar von Cope wurde mit gesenktem Kopf auf allen vier Beinen dargestellt, als würde es fressen, da sein Schädel besonders gut erhalten war. Das von Brown geborgene Skelett erhielt dagegen eine zweibeinige Haltung mit erhobenem Kopf, weil sein Schädel unvollständiger war. Henry F. Osborn erklärte diese Szene als Darstellung zweier Tiere, die gemeinsam in einem sumpfigen Gebiet Nahrung aufnehmen, während das aufgerichtete Individuum durch die Annäherung eines Tyrannosaurus aufgeschreckt wird. Zur Gestaltung des Dioramas wurden außerdem Pflanzen- und Muschelreste ergänzt, die auf tatsächlich gemeinsam gefundenen Fossilien basierten, darunter Ginkgoblätter, Zapfen von Mammutbäumen und Schachtelhalme.

Aufgrund der damals noch sehr spärlichen Erkenntnisse über Hadrosaurier kam es in den folgenden Jahrzehnten immer wieder zu unterschiedlichen Einordnungen der beiden Exemplare. Lehrbücher und Enzyklopädien unterschieden häufig zwischen dem „Iguanodon-ähnlichen“ Claosaurus annectens und dem „entenschnäbligen“ Hadrosaurus, dessen Fossilien heute als erwachsene Exemplare von Edmontosaurus annectens erkannt werden. John B. Hatcher vertrat dagegen ausdrücklich die Ansicht, dass C. annectens mit genau diesem entenschnäbligen Hadrosaurier identisch sei. In seiner 1902 veröffentlichten Revision nahm Hatcher weitreichende Änderungen vor und betrachtete nahezu alle damals bekannten Hadrosaurier-Gattungen als Synonyme von Trachodon. Dazu zählten neben Claosaurus auch die Gattungen Cionodon, Diclonius, Hadrosaurus, Ornithotarsus, Pteropelyx und Thespesius sowie die fragmentarisch bekannten Gattungen Claorhynchus und Polyonax, die heute als Ceratopsier angesehen werden.20Hatcher, J. B. (1902). The genera and species of the Trachodontidae (Hadrosauridae, Claosauridae) Marsh. Annals of the Carnegie Museum, 1(14), 377–386. doi:10.5962/p.331063 Hatchers Arbeit führte zunächst zu einem breiten wissenschaftlichen Konsens, und so trug der Dinosaurier, um den es hier weiterhin geht, nun für lange Zeit den Namen Trachodon annectens.
Thespesius
 Charles W. Gilmore Charles W. Gilmore |
Nach 1910 wurden jedoch neue Fossilien aus Kanada und Montana entdeckt, die eine wesentlich größere Vielfalt der Hadrosaurier erkennen ließen als zuvor angenommen. Charles W. Gilmore, der einen großen Teil von Marshs umfangreicher Fossiliensammlung nach dessen Tod 1899 verwaltete, überprüfte die Gruppe im Jahre 1915 erneut. Er schlug vor, den Gattungsnamen Thespesius für Hadrosaurier aus der Lance Formation und gleichaltrigen Gesteinseinheiten wieder einzuführen. Den Namen Trachodon wollte er dagegen auf einen Hadrosaurier aus der älteren Judith River Formation und vergleichbaren Schichten beschränken, da dessen Typusmaterial nur unzureichend war. Bezüglich Claosaurus annectens schlug Gilmore vor, diese Art mit Thespesius occidentalis gleichzusetzen.21Gilmore, C. W. (1915). On the genus Trachodon. Science, 41(1061), 658–660. doi:10.1126/science.41.1061.658 |
 Ferdinand V. Hayden Ferdinand V. Hayden |
Der Name „Thespesius“ hat sogar eine noch längere und mindestens ebenso verwirrende Geschichte. Schon im Jahr 1855 hatte der Geologe Ferdinand V. Hayden mehrere Fossilien an den Paläontologen Joseph Leidy nach Philadelphia geschicht, die aus Schichten stammten, die damals als Great Lignite Formation bezeichnet wurden. Heute gelten sie als Teil der Hell Creek Formation. Der Fundort lag im damaligen Nebraska-Territorium nahe dem Grand River im Gebiet des heutigen South Dakota. Zu den Fossilien gehörten zwei Schwanzwirbel mit den Sammlungsnummern USNM 219 und USNM 221 sowie eine mittlere Zehenphalanx, die mit der Nummer USNM 220 inventarisiert wurde. |
| Joseph Leidy |
Zwei Jahre später, also 1856, benannte Joseph Leidy auf Grundlage dieser drei Knochen die Typusart Thespesius occidentalis. Der Gattungsname leitet sich vom griechischen Wort θεσπέσιος (thespesios, „wunderbar“ oder „erstaunlich“) ab und bezieht sich auf die gewaltige Größe der Überreste. Der Artname occidentalis bedeutet auf Latein „westlich“. Leidy verzichtete beim Gattungsnamen bewusst auf die Endung „saurus“, weil Hayden angegeben hatte, die Fossilien stammten aus einer miozänen Gesteinsschicht und sollten somit wohl eher zu einem Säugetier gehören als zu einem Dinosaurier. Leidy selbst war jedoch überzeugt, dass es sich um einen Dinosaurier handelte.22Leidy, J. (1856). Notice of extinct Vertebrata, discovered by F. V. Hayden during the expedition to the Sioux country under the command of Lieut. G. K. Warren. Proceedings of the Academy of Natural Science Philadelphia, 8, 311–312. Es ist sehr wahrscheinlich, dass es sich dabei auch um Überreste von dem Tier handelt, das wir heute Edmontosaurus annectens nennen. |
Schon lange bevor Charles W. Gilmore den Namen „Thespesius“ wieder ins Spiel brachte, war er von einer ganzen Reihe von Autoren immer wieder als Sammelbezeichnung für nahezu alle Hadrosaurier aus dem späten Maastrichtium Nordamerikas verwendet worden.23Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210. 1875 hatte z.B. Edward D. Cope plausibel dargelegt, dass er Agathaumas milo, das nur auf unvollständigen Gliedmaßenknochen und einigen Wirbeln beruhte, für ein Synonym von Thespesius occidentalis halte, welches er damals selbst als Art von Hadrosaurus ansah.24Cope, E. D. (1875). Report on the vertebrate paleontology of Colorado. US Government Printing Office. Im gleichen Jahr, in welchem Marsh seinen Claosaurus beschrieb, veröffentlichte er auch den Namen Hadrosaurus longiceps und stellte das Exemplar somit in eine Gattung, die Joseph Leidy bereits 1858 auf Basis von Fossilien aus New Jersey beschrieben hatte.25Leidy, J. (1858). Hadrosaurus foulkii, a new saurian from the Cretaceous of New Jersey, related to Iguanodon. Proceedings of the Academy of Natural Sciences of Philadelphia, 10, 213–218.
 Frederic A. Lucas Frederic A. Lucas |
Im Jahr 1900 veröffentlichte Frederic A. Lucas einen kurzen Beitrag in der Fachzeitschrift Science, in dem er darauf hinwies, dass sich die ursprünglichen Fossilien von Thespesius occidentalis nicht von vollständigeren Exemplaren unterscheiden ließen, die im späten 19. Jahrhundert als Claosaurus annectens bezeichnet worden waren. Daher argumentierte Lucas, dass für dieses Tier der ältere Name Thespesius occidentalis verwendet werden sollte.26Lucas, F. A. (1900). Paleontological Notes. Science, 12(308), 809–810. |
Auf dieser Grundlage stützte 1915 auch Gilmore seine Argumentation. Er stellte außerdem fest, dass die Typusfossilien von Thespesius zwar unzureichend seien, geologische Untersuchungen aber eindeutig zeigten, dass sie aus denselben Schichten stammten wie Claosaurus bzw. Trachodon, wie zum Beispiel auch die Exemplare, die Edward D.Cope während der Bone Wars entdeckt hatte und die im American Museum of Natural History montiert und ausgestellt worden waren. Deshalb sollte seiner Ansicht nach der ältere Name T. occidentalis für die Hadrosaurier der Lance Formation verwendet werden.27Gilmore, C. W. (1915). On the genus Trachodon. Science, 41(1061), 658–660. doi:10.1126/science.41.1061.658 Gilmore beschrieb 1924 auch selbst eine eigene Art, Thespesius edmontoni, auf Basis von Fossilien aus dem kanadischen Alberta.28Gilmore, C. W. (1924). A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta. Canada Department of Mines Geological Survey Bulletin (Geological Series), 38(43), 13–26.
 Loris S. Russel Loris S. Russel
|
Viele spätere Forscher, darunter Loris S. Russell29Russell, L. S. (1930). Upper Cretaceous dinosaur faunas of North America. Proceedings of the American Philosophical Society, 69(1), 133–159. und Charles M. Sternberg30Sternberg, C. M. (1936). The systematic position of Trachodon. Journal of Paleontology, 10(7), 652–655.31Sternberg, C. M. (1939). Were there Proboscis-bearing Dinosaurs? Discussion of Cranial Protuberances in the Hadrosauridae. Journal of Natural History, 3(17), 556–560., folgten dieser Einschätzung. Sie verwendeten in den 1920er- und 1930er-Jahren weiterhin die Bezeichnungen Thespesius occidentalis, allerdings auch hin und wieder Thespesius annectens für Hadrosaurier aus der Lance Formation. Sternberg beschrieb 1926 noch eine weitere Art, Thespesius saskatchewanensis auf Basis eines großen Unterkiefers (Inventarnummer NMC 8509), der in der kanadischen Frenchman Formation gefunden und 1890 von Othniel C. Marsh 1897 als Trachodon longiceps beschrieben worden war.32Marsh, O. C. (1897). Vertebrate fossils of the Denver Basin. U.S. Geological Survey, Monthly, 27, 473–527.33Sternberg, C. M. (1926). A new species of Thespesius from the Lance Formation of Saskatchewan. Canada Department of Mines Geological Survey Bulletin (Geological Series), 44(46), 73–84. |

Edmontosaurus
 Levi Sternberg Levi Sternberg
|
Die Familie Sternberg, insbesondere Charles Brüder Levi und George, waren bereits zuvor mit den Entenschnabeldinosauriern eng verbunden. Ein weiteres Exemplar mit der Inventarnummer NMC 2288 besteht aus einem Schädel, der Wirbelsäule bis zum sechsten Schwanzwirbel, Rippen, Teilen des Beckens, einem Oberarmknochen sowie dem Großteil eines Hinterbeins. Es wurde 1912 von Levi Sternberg in der Horseshoe Canyon Formation (früher als untere Edmonton Formation bezeichnet) entlang des Red Deer River im südlichen Alberta entdeckt. Dort kam noch ein weiterer Fund zum Vorschein, mit Schädel und ein Skelett, dem jedoch der Schnabel, der größte Teil des Schwanzes und Teile der Füße fehlen. Dieses Exemplar wurde 1916 von Levis Bruder George F. Sternberg gefunden. |

 Lawrence M. Lambe Lawrence M. Lambe |
Beide Exemplare wurden in die Sammlung des Geological Survey of Canada in Ottawa überführt und dort 1917 von Lawrence M. Lambe weiter untersucht. Lambe stellte zwar fest, dass sein neuer Dinosaurier den Exemplaren von Diclonius mirabilis sehr ähnlich war, hob jedoch die deutlich größere Körpergröße und den kräftigeren Körperbau der neuen Exemplare hervor, weshalb er ihnen einen eigenen Namen gab: Edmontosaurus regalis. Der Gattungsname nimmt einerseits Bezug auf die Edmonton Formation, die wiederum ihren Namen der Hauptstadt des Bundesstaates Alberta, nämlich Edmonton verdankt. Der Artname bedeutet soviel wie „königlich“ oder „von königlicher Größe“.34Lambe, L. M. (1917). A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta. The Ottawa Naturalist, 31(7), 65–73. |
Lambe hatte bereits 1902 eine weitere Hadrosaurier-Art als Trachodon selwyni anhand eines Unterkiefers aus der heute als Dinosaur Park Formation bezeichneten Gesteinseinheit in Alberta beschrieben.35Lambe, L. M. (1902). On Vertebrata of the mid-Cretaceous of the Northwest Territory. New genera and species from the Belly River Series (mid-Cretaceous). Contributions to Canadian Paleontology, 3, 25–81. Diese gilt heute aber auch als zweifelhaft und dürfte mit Edmontosaurus regalis übereinstimmen.36Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272. 1920 veröffentlichte Lambe auch eine Beschreibung des Schädels von NMC 2289.37Lambe, L. M. (1920). The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta. Memoir, 120. Department of Mines, Geological Survey of Canada, 1–79. Doch der größte Teil des Skeletts ist bis heute nicht ausführlich wissenschaftlich beschrieben worden und befindet sich noch immer in seinen Gipsmanschetten.
Die ersten bekannten Fossilien, die sehr wahrscheinlich zu Edmontosaurus regalis gehören, wurden aber bereits 1871 von Edward D. Cope unter dem Namen Trachodon cavatus beschrieben.38Cope, E. D. (1871). Supplement to the synopsis of the extinct Batrachia and Reptilia of North America. Proceedings of the American Philosophical Society, 12(86), 41–52. In manchen Quellen wird dieser Name aber auch häufig als T. atavus39Glut, D. F. (1997). Edmontosaurus. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 389–396. oder auch als T. atavus wiedergegeben.40Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 438–463.41Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1. Aufl.). University of California Press, Berkeley, 534–561. Dies ist wahrscheinlich ein Resultat der Bone Wars, die sich in diesem Jahr auf einem ersten Höhepunkt befanden. Cope und Marsh brachten in dieser Zeit in hohem Tempo mit ihren neuen Benennungen die Telegrafenstationen regelrecht zum glühen, um den jeweils anderen einerseits bei der Beschreibung neuer Dinosaurier zu übertreffen. Beide vergaben oft mehrere Namen für ein und dasselbe Tier, was später als „wissenschaftliches Flächenbombardement“ bezeichnet wurde. Außerdem passierten an den Telegrafiestationen aufgrund dieser Praxis verständlicherweise jede Menge Fehler, was bei der Wortähnlichkeit hier ebenfalls sehr wahrscheinlich ist. 1874 führte Cope außerdem den Namen Agathaumas milo für ein Kreuzbeinwirbel-Fragment und Teile eines Schienbeins aus der oberkreidezeitlichen Laramie Formation in Colorado ein, beschrieb die Fossilien jedoch zunächst nicht näher.42Cope, E. D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. U.S. Geological and Geographical Survey of the Territories Annual Report, 1, 9–28. Noch im selben Jahr veröffentlichte er eine Beschreibung derselben Knochen unter dem Namen Hadrosaurus occidentalis.43Cope, E. D. (1874). Report on the vertebrate paleontology of Colorado. U.S. Geological and Geographical Survey of the Territories Annual Report, 2, 429–454. Bedauerlicherweise gelten diese Fossilien heute aber alle als verschollen.44Carpenter, K. & Young, D. B. (2002). Late Cretaceous dinosaurs from the Denver Basin, Colorado. Rocky Mountain Geology, 37(2), 237–254. doi:10.2113/11 Obwohl Trachodon atavus mehrere Jahrzehnte früher benannt worden war als E. regalis, stuften spätete Arbeiten diese Art ohne Kommentar als Synonym von Edmontosaurus regalis ein, genauso auch Agathaumas milo.45Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 438–463.46Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1. Aufl.). University of California Press, Berkeley, 534–561. Sie waren in den kommenden Jahren überhaupt nicht mehr verwendet wurden und hatten auch sonst in der Forschung kaum Beachtung gefunden, was sicherlich auch ein Resultat der Bone Wars ist.
Anatosaurus
Bereits vor Gilmores weitreichender Argumentation hatte Lawrence Lambe jedoch die Ansicht vertreten, dass die Typusfossilien von Thespesius occidentalis zu unvollständig seien und daraus gezogene Schlussfolgerungen ebenso spekulativ wären wie im Fall von Trachodon, wie er in einer Arbeit im Jahre 1913 darlegte.
 Richard S. Lull Richard S. Lull
|
1942 versuchten Richard S. Lull und Nelda E. Wright, die komplizierte Taxonomie der Hadrosaurier, insbesondere derer, von denen noch kein Schädel vorlag, zu vereinfachen. Dazu führten sie die neue Gattung Anatosaurus ein. Der Name ist ein lateinisch-griechisches Mischwort und bedeutet „Enten-Echse“. Er bezieht sich auf den breiten, entenähnlichen Schnabel. Als Typusart bestimmten sie Marshs frühere Art Claosaurus annectens, die ja damals eigentlich schon Thespesius hieß. Mit Anatosaurus bekam der Dinosaurier folglich seinen dritten Namen. Zusätzlich ordneten sie auch Thespesius edmontoni und T. saskatchewanensis dieser Gattung zu. Eine neue Art beschrieben sie auch selbst: Anatosaurus copei beruhte auf zwei Skeletten im American Museum of Natural History, die Cope als Diclonius mirabilis beschrieben hatte. So entstanden also die Arten Anatosaurus annectens, A. copei, A. edmontoni, A. longiceps und A. saskatchewanensis.47Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272.
Obwohl sie in ihrer Arbeit einräumten, dass sich Thespesius occidentalis möglicherweise anhand seiner kürzeren Schwanzwirbel von Anatosaurus unterscheiden lasse, schlossen sie sich letztlich Lambes Einschätzung an, dass Thespesius occidentalis trotz seiner historischen Bedeutung zu unvollständig für einen zuverlässigen Vergleich sei. Seitdem wird die Gattung Thespesius allgemein als nomen dubium betrachtet und weitgehend ignoriert. Der Name Anatosaurus wurde in der Folge aber zu dem wohl bekanntesten Namen für einen Hadrosaurier und noch bis zum Ende des 20. Jahrhunderts als der „klassische Entenschnabeldinosaurier“ schlechthin bezeichnet.48Glut, D. F. (1982). The New Dinosaur Dictionary. Citadel Press, Secaucus, NJ, 57. |

Anatotitan
 Michael K. Brett-Surman Michael K. Brett-Surman |
Diese Einteilung blieb über mehrere Jahrzehnte bestehen, bis Michael K. Brett-Surman das betreffende Fossilmaterial im Rahmen seiner Abschlussarbeiten in den 1970er- und 1980er-Jahren erneut untersuchte. Er kam zu dem Schluss, dass die Typusart Anatosaurus annectens tatsächlich zu Edmontosaurus gehört und dass sich A. copei deutlich genug unterscheidet, um eine eigene Gattung zu rechtfertigen.49Brett-Surman, M. K. (1975). The appendicular anatomy of hadrosaurian dinosaurs. M.A. thesis, University of California, Berkeley.50Brett-Surman, M. K. (1979). Phylogeny and paleobiogeography of hadrosaurian dinosaurs. Nature, 277(5697), 560–562. doi:10.1038/277560a051Brett-Surman, M. K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation, George Washington University, Washington, D.C. |
Obwohl Dissertationen nach den Regeln des International Code of Zoological Nomenclature (ICZN) nicht als offizielle Veröffentlichungen gelten, wurden Brett-Surmans Ergebnisse von anderen Paläontologen bekannt gemacht und in mehreren populärwissenschaftlichen Werken übernommen.52Glut, D. F. (1982). The New Dinosaur Dictionary. Citadel Press, Secaucus, NJ, 49, 53.53Lambert, D. & the Diagram Group (1983). A Field Guide to Dinosaurs. Avon Books, New York, 156–161.
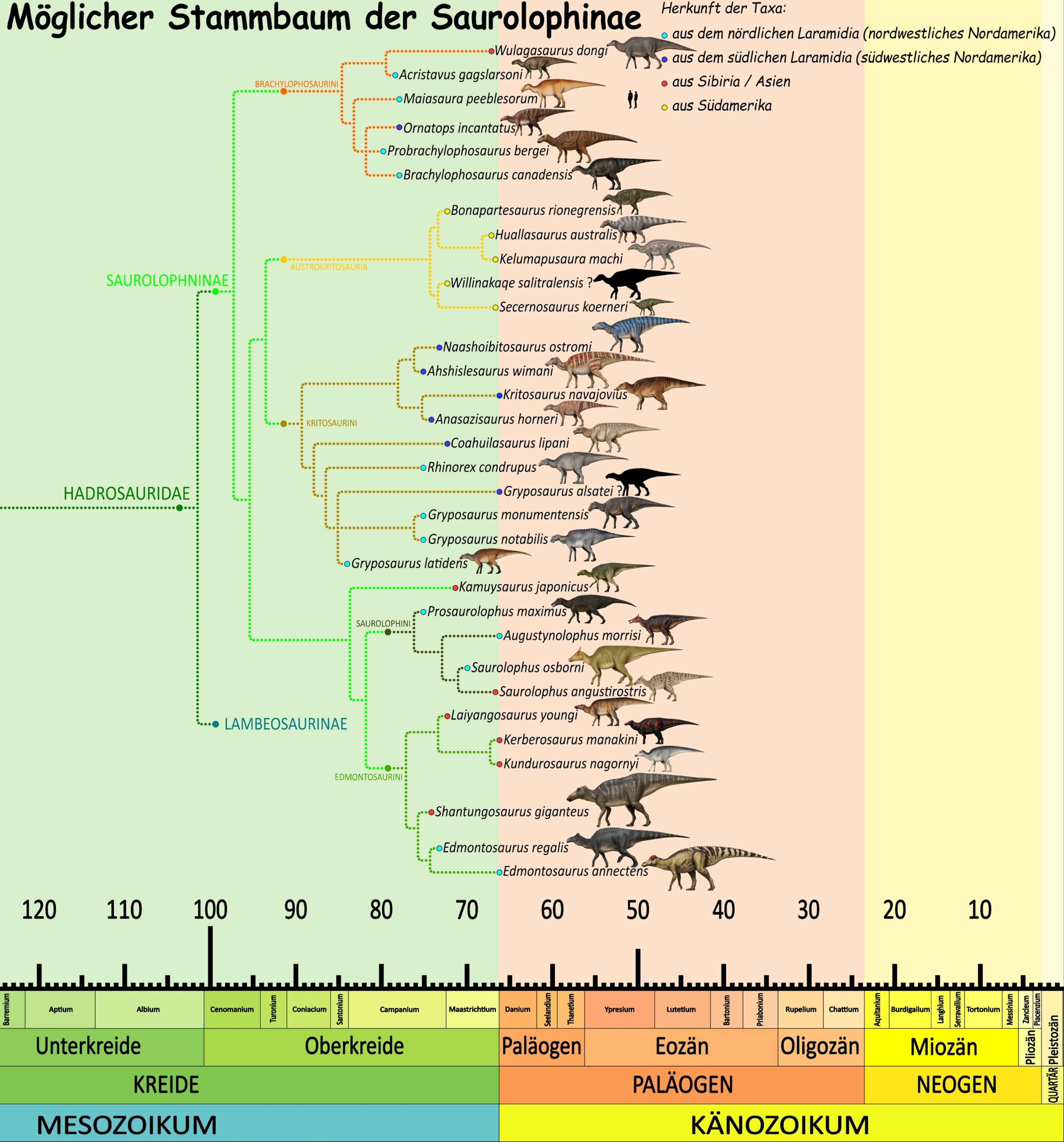
1990 stellten Michael K. Brett-Surman und Ralph Chapman deshalb für A. copei die neue Gattung Anatotitan auf.54Chapman, R. E. & Brett-Surman, M. K. (1990). Morphometric observations on hadrosaurid ornithopods. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Perspectives and Approaches. Cambridge University Press, Cambridge, 163–177. Die übrigen Arten wurden ebenfalls neu eingeordnet: Anatosaurus saskatchewanensis und Anatosaurus edmontoni wiesen sie der Gattung Edmontosaurus zu. Anatosaurus longiceps wurde wiederum Anatotitan zugeordnet, entweder als eigenständige Art oder als Synonym von A. copei. Da die Typusart von Anatosaurus (A. annectens) nun zu Edmontosaurus gestellt wurde, gilt der Name Anatosaurus seither als jüngeres Synonym (junior synonym) von Edmontosaurus und sollte nicht mehr verwendet werden.55Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1st ed.). University of California Press, Berkeley, 534–561.56Olshevsky, G. (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. Mesozoic Meanderings, 2. Publications Requiring Research, San Diego. Das daraus entstandene Gattungskonzept von Edmontosaurus umfasste zunächst also drei gültige Arten: E. regalis als Typusart, E. annectens (einschließlich Anatosaurus edmontoni, später zu edmontonensis korrigiert), sowie E. saskatchewanensis.
Neuere Diskussionen
Die Diskussion über die korrekte taxonomische Stellung der Exemplare von Anatotitan copei-Exemplare hält bis heute an. In Anlehnung an die bereits 1902 von John B. Hatcher vertretene Auffassung betrachteten Jack Horner, David B. Weishampel und Catherine Forster Anatotitan copei als Exemplare von Edmontosaurus annectens, deren Schädel während der Fossilisation zusammengedrückt worden waren.57Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2nd ed.). University of California Press, Berkeley, 438–463.
2007 wurde ein weiteres außergewöhnlich gut erhaltenes Fossil, eine sogenannte Mumie, bekannt gegeben. Das Exemplar erhielt den Spitznamen „Dakota“ und die Inventarnummer NDGS 2000. Es war bereits 1999 von Tyler Lyson entdeckt worden und stammt aus der Hell Creek Formation in North Dakota.58Lee, C. (2007). Scientists Get Rare Look at Dinosaur Soft Tissue. The Washington Post. In einer 2011 veröffentlichten Studie führten Nicolás Campione und David Evans erstmals eine morphometrische Analyse an diesem Exemplar durch, bei der die verschiedenen Edmontosaurus-Exemplare quantitativ miteinander verglichen wurden. Sie kamen zu dem Ergebnis, dass nur zwei Arten als gültig anzusehen sind: Edmontosaurus regalisaus dem späten Campanium und Edmontosaurus annectensaus dem späten Maastrichtium. Darüber hinaus lieferten ihre Untersuchungen weitere Belege dafür, dass Anatotitan copei lediglich ein Synonym von Edmontosaurus annectens ist. Insbesondere erklärten sie den langen, flachen Schädel von A. copei als Folge ontogenetischer Veränderungen und interpretierten ihn als Merkmal vollständig ausgewachsener Individuen von E. annectens.59Campione, N. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186
Eine Studie von Nicolás Campione und David Evans aus dem Jahr 2011 kam zu dem Ergebnis, dass es sich beim von Charles M. Sternberg beschriebenen Thespesius saskatchewanensis um ein Synonym von Edmontosaurus annectens handelt, während Thespesius edmontoni von Charles W. Gilmore ein Synonym von Edmontosaurus regalis ist.60Campione, N. S. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186

Heute wird Edmontosaurus annectens von Edmontosaurus regalis vor allem durch seinen längeren, flacheren und insgesamt weniger kräftig gebauten Schädel unterschieden. Michael K. Brett-Surman vermutete zwar, dass E. regalis und E. annectens möglicherweise männliche und weibliche Vertreter derselben Art sein könnten, doch stammen sämtliche bekannten Fossilien von E. regalis aus älteren Gesteinsschichten als die von E. annectens. E. regalis lebte bereits im Campanium bis ins frühe Maastrichtium, also in der Zeit vor 73 bis 70 Millionen Jahren. E. annectens kommt dagegen nur im späten Maastrichtium, in der Zeit vor 68 bis 66 Ma vor. 61Glut, D. F. (1997). Anatotitan. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 132–134.62Campione, N. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.002518663Brett-Surman, M. K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation, George Washington University, Washington, D.C. Es ist somit durchaus möglich, dass beide Arten direkt miteinander verbundene Chronospezies waren, also dass E. annectens ein direkter Nachfahre von E. regalis ist.



Beschreibung
Der Schädel und das Skelett des Tieres, das wir heute Edmontosaurus annectens nennen, sind außergewöhnlich gut bekannt. Es handelt sich um einen sehr großen Dinosaurier, einen der größten überhaupt seines damaligen Lebensraums. Bereits Edward D. Cope schätzte die Länge des ersten gefundenen Exemplars ursprünglich auf etwa 12 Meter, mit einer Schädellänge von etwa 1,18 Metern.76Cope, E. D. (1883). On the characters of the skull in the Hadrosauridae. Proceedings of the Philadelphia Academy of Natural Sciences, 35, 97–107. Diese Schätzung wurde später zwar nach unten korrigiert, heute geht man für dasselbe Exemplar von einer Gesamtlänge von etwa 8,8 Metern aus.77Lull, R. S. & Wright, N. E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper, 40. Geological Society of America, 225. Zu Copes Verteidigung muss man allerdings sagen, dass der Bach, wo das Fossil gefunden wurde, bereits ein gutes Dutzend Wirbel, das Becken, die Oberschenkelknochen und den hinteren Teil des Schwanzes fortgespült hatte.78Osborn, H. F. (1909). The Upper Cretaceous iguanodont dinosaurs. Nature, 81(2075), 160–162. doi:10.1038/081160a0 Außerdem sind heute tatsächlich Exemplare bekannt, die auf eine Körperlänge von 12 Metern und sogar mehr schließen lassen.79Morris, W. J. (1970). Hadrosaurian dinosaur bills. Morphology and function. Contributions in Science (Los Angeles County Museum of Natural History), 193, 1–14.80Sues, H.-D. (1997). Ornithopods. In Farlow, J. O. & Brett-Surman, M. K. (Hgg.), The Complete Dinosaur. Indiana University Press, Bloomington, 338. Ein ausgewachsener Edmontosaurus annectens wog vermutlich zwischen 6 und 8 Tonnen.81Glut, D. F. (1997). Anatotitan. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 132–134.
Einige neuere Exemplare, die derzeit noch im Museum of the Rockies in Bozeman in Montana untersucht werden, weisen auf noch deutlich größere Giganten hin. „X-rex“ (MOR 1142) und „Becky’s Giant“ (MOR 1609) deuten darauf hin, dass E. annectens möglicherweise Längen von 15 Metern erreichen konnte, mit einem Gewicht von 15,87 Tonnen. Solche Individuen dürften jedoch eher selten gewesen sein.82Horner, J. R., Goodwin, M. B. & Myhrvold, N. (2011). Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLOS ONE, 6(2), e16574. doi:10.1371/journal.pone.001657483Prieto-Márquez, A. (2014). A juvenile Edmontosaurus from the late Maastrichtian (Cretaceous) of North America. Implications for ontogeny and phylogenetic inference in saurolophine dinosaurs. Cretaceous Research, 50, 282–303. doi:10.1016/j.cretres.2014.05.00384Jiménez-Moreno, F. J. et al. (2025). First Population Analysis in Hadrosauroid dinosaurs (Ornithopoda: Iguanodontia: Hadrosauroidea). Evolving Earth, 3, 100072. doi:10.1016/j.eve.2025.100072 Eine Studie aus dem Jahr 2022 zur Knochenhistologie und zum Wachstum von E. annectens kam zu dem Schluss, dass frühere Größenschätzungen teilweise auch deutlich zu hoch, aber auch zu niedrig ausgefallen sein könnten. Nach dieser Untersuchung erreichten vollständig ausgewachsene Tiere gewöhnlich eine Länge von etwa 11 bis 12 Metern und ein durchschnittliches Körpergewicht von 5,6 bis 7 Tonnen. Damit waren sie ungefähr so massig wie ein heutiger afrikanischer Elefant.
In Die Weißen Steine bin ich von diesen eher moderaten Rekonstruktionen ausgegangen, habe aber die Möglichkeit von sehr großen Ausnahme-Exemplaren durchaus mit einbezogen. In der Neuauflange von Band I werden John und Razor von einem riesigen Edmontosaurus angegriffen, der über 14 Meter lang ist.85Wosik, M. et al. (2022). Osteohistological and taphonomic life-history assessment of Edmontosaurus annectens (Ornithischia: Hadrosauridae) from the Late Cretaceous (Maastrichtian) Ruth Mason dinosaur quarry, South Dakota, United States, with implication for ontogenetic segregation between juvenile and adult hadrosaurids. Journal of Anatomy, 241(2), 272–296. doi:10.1111/joa.13679
Bau des Schädels und mögliche Ernährungsweise
Der Schädel von Edmontosaurus annectens ist vor allem für seine lange und breite Schnauze bekannt. Im Verhältnis zum übrigen Körper war der Schnabel länger und flacher als bei jedem anderen bekannten Hadrosaurier. Auch der zahnlose Vorderabschnitt des Unterkiefers war im Verhältnis länger als bei allen anderen Vertretern dieser Dinosauriergruppe.86Chapman, R. E. & Brett-Surman, M. K. (1990). Morphometric observations on hadrosaurid ornithopods. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Perspectives and Approaches. Cambridge University Press, Cambridge, 163–177. Edward D. Cope verglich ihn in seiner Erstbeschreibung in der Seitenansicht mit der eines Gänseschnabels und in der Draufsicht mit dem kurzen Schnabel eines Löfflers. Cope ging seiner Zeit davon aus, dass Hadrosaurier überwiegend im Wasser gelebt hätten. Er begründete dies damit, dass die Zähne des Unterkiefers seiner Meinung nach nur locker im Knochen verankert gewesen seien und beim Fressen harter Landpflanzen leicht hätten ausbrechen können. Auch den Schnabel hielt er für wenig stabil.87Cope, E. D. (1883). On the characters of the skull in the Hadrosauridae. Proceedings of the Philadelphia Academy of Natural Sciences, 35, 97–107. Auch William Morris vermutete, dass sich das Tier ähnlich wie einige heutige Enten ernährte, indem es Pflanzen sowie kleine Wassertiere wie Weichtiere und Krebstiere aus dem Wasser filterte und das überschüssige Wasser über V-förmige Rinnen an der Innenseite des Oberschnabels wieder ausströmen ließ.88Morris, W. J. (1970). Hadrosaurian dinosaur bills. Morphology and function. Contributions in Science (Los Angeles County Museum of Natural History), 193, 1–14.
Diese Einschätzung beruhte jedoch auf einer Fehlinterpretation. Zufällig fehlten Copes Fossil nämlich die inneren Knochenwände des Unterkiefers, welche die Zähne normalerweise stützten. Bei anderen Exemplaren waren diese aber durchaus erhalten. Tatsächlich waren die Zähne somit sehr fest im Kiefer verankert und sogar ausgesprochen gut zum Kauen geeignet.89Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272.90Ostrom, J. H. (1964). A reconsideration of the paleoecology of the hadrosaurian dinosaurs. American Journal of Science, 262(8), 975–997. doi:10.2475/ajs.262.8.975 Die Rinnen und Leisten des Schnabels ähneln zudem vielmehr den Schnäbeln pflanzenfressender Schildkröten als den flexiblen Filterstrukturen moderner filtrierender Vögel.91Barrett, P. M. (2005). The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria). Palaeontology, 48(2), 347–358. doi:10.1111/j.1475-4983.2005.00448.x Da Kratzspuren die mikroskopischen Abnutzungsspuren auf den Zähnen dominieren, ist es wahrscheinlich, dass Edmontosaurus eher bodennahe Pflanzen und nicht so sehr ein selektiver Blattfresser gewesen sein könnte. Als Ursache für die stärkere Zahnabnutzung kommen kieselsäurereiche Pflanzen wie Schachtelhalme oder versehentlich mit der Nahrung aufgenommene Bodenpartikel infrage, da Edmontosaurus häufig in Bodennähe fraß.92Williams, V. S., Barrett, P. M. & Purnell, M. A. (2009). Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. Proceedings of the National Academy of Sciences, 106(27), 11194–11199. doi:10.1073/pnas.0812631106 Vergleiche der mikroskopischen Zahnabnutzung mit anderen Hadrosauriern zeigen jedoch, dass die Nahrung von Edmontosaurus wahrscheinlich nicht wesentlich härter war als die seiner nahen Verwandten.93Kubo, T. et al. (2023). Dental microwear texture analysis reveals a likely dietary shift within Late Cretaceous ornithopod dinosaurs. Palaeontology, 66(6), e12681. doi:10.1111/pala.12681 In höheren Breiten, besonders in Alaska, dürften Nadelbäume ebenfalls eine wichtige Nahrungsquelle für ihn dargestellt haben. Die Zusammensetzung der Nahrung änderte sich dort vermutlich saisonal im Verlauf des Jahres.94Fiorillo, A. R. (2011). Microwear patterns on the teeth of northern high latitude hadrosaurs with comments on microwear patterns in hadrosaurs as a function of latitude and seasonal ecological constraints. Palaeontologia Electronica, 14(3), 1–17.

Der Bau der Zähne deutet jedenfalls darauf hin, dass Edmontosaurus sowohl seine aufgenommene Nahrung zerschneiden als auch mahlend kauen konnte.95Erickson, G. M. et al. (2012). Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science, 338(6103), 98–101. doi:10.1126/science.122449596Rybczynski, N. et al. (2008). A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses. Palaeontologia Electronica, 11(2), online publication.97Williams, V. S., Barrett, P. M. & Purnell, M. A. (2009). Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. Proceedings of the National Academy of Sciences, 106(27), 11194–11199. doi:10.1073/pnas.0812631106 Eine 2008 veröffentlichte Studie von Casey Holliday und Lawrence Witmer kam zu dem Ergebnis, dass Ornithopoden wie Edmontosaurus allerdings nicht über die beweglichen Schädelgelenke verfügten, die man bei heutigen Echsen oder Vögeln mit beweglichem Schädel findet. Sie stattdessen vor, dass einige Strukturen, die zuvor als bewegliche Gelenke interpretiert worden waren, in Wirklichkeit knorpelige Wachstumszonen gewesen seien. Allein durch Kratzspuren auf den Zähnen könne sich daher nicht die Bewegungsrichtung des Unterkiefers ableiten lassen. Ähnliche Spuren können auch durch andere Bewegungen entstehen, etwa durch geringe Verschiebungen zwischen den beiden Unterkieferhälften.98Holliday, C. M. & Witmer, L. M. (2008). Cranial kinesis in dinosaurs. Intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids. Journal of Vertebrate Paleontology, 28(4), 1073–1088. doi:10.1671/0272-4634-28.4.1073 Wie genau Hadrosaurier ihre Nahrung zerkaut haben könnten, wurde auch bereits sowohl mit Laserscans, Computersimulationen und aufwendigeren dreidimensionalen Animationsmodellen erforscht. Dabei zeigte sich, dass sich die Knochen um maximal 1,3 bis 1,4 Zentimeter voneinander entfernen konnten.99Rybczynski, N. et al. (2008). A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses. Palaeontologia Electronica, 11(2), online publication.100Williams, V. S., Barrett, P. M. & Purnell, M. A. (2009). Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. Proceedings of the National Academy of Sciences, 106(27), 11194–11199. doi:10.1073/pnas.0812631106 Die Mahlbewegung wurde wahrscheinlich vor allem durch den Unterkiefer erzeugt. Dieser konnte sich nicht nur wie üblich auf- und abbewegen, sondern auch leicht nach vorne und hinten verschieben. Zusätzlich erlaubte das Gelenk zwischen den beiden Unterkieferhälften geringe Vor- und Rückbewegungen sowie eine leichte Drehung. Diese Kombination würde die beobachteten Zahnabnutzungsspuren erklären und passt gleichzeitig zu dem insgesamt sehr stabil gebauten Schädel von Edmontosaurus.101Cuthbertson, R. S. et al. (2012). Kinetic limitations of intracranial joints in Brachylophosaurus canadensis and Edmontosaurus regalis (Dinosauria: Hadrosauridae), and their implications for the chewing mechanics of hadrosaurids. The Anatomical Record, 295(6), 968–979. doi:10.1002/ar.22458
Berichte über sogenannte Gastrolithen, also Magensteine, bei Edmontosaurus beruhen vermutlich auf einer Fehlinterpretation. Es handelt sich wohl lediglich um Kiesel, die erst während der Einbettung des Kadavers in das Sediment in den Körper gelangten.102Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.
Ein erhaltenes Hornschnabelmaterial (Rhamphotheca) des Exemplars LACM 23502, das im Los Angeles County Museum aufbewahrt wird, zeigt, dass der Schnabel hakenförmig gebogen und breiter war, als er in vielen wissenschaftlichen Rekonstruktionen und populären Darstellungen bislang gezeigt wurde.103Naish, D. (2018). Enough with the Duck-Billed Dinosaurs. Scientific American.104Black, R. (2012). Shovel-Beaked, Not Duck-Billed. Smithsonian Magazine. Damitl rupfte Edmontosaurus Pflanzen ab, während wangähnliche Weichteilstrukturen die Nahrung beim Kauen im Maul hielten. Sein hochentwickelter Schädel ermöglichte eine mahlende Kaubewegung, die funktionell dem Kauen heutiger Säugetiere ähnelte. Er besaß hunderte Zähne, die wie bei den meisten Hadrosauriern eine Art Batterie bildeten. Insgesamt befanden sich im Maul eines Edmontosaurus zwischen 1.000 und 1.500 Zähne gleichzeitig. Etwa 200 bis 300 davon waren als Kaufläche angeordnet. Für jeden einzelnen standen aber bereits mehrere als Reserve bereit, wenn der Zahn abgekaut war und ausfiel. So wurden die Zähne während des gesamten Lebens ständig ersetzt. Die Nahrungssuche erfolgte vermutlich vom Boden bis in Höhen von etwa 4 Metern, sodass sowohl niedrige als auch höher wachsende Pflanzen erreicht werden konnten.105Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2nd ed.). University of California Press, Berkeley, 438–463.






Haut und Weichgewebe
Durch die vielen ausgezeichnet erhaltenen Fossilmumien wie „Edmond“ (SM 4036) und „Dakota“ (NDGS 2000) sind uns auch umfassende Hautpartien überliefert, sowie die Hornscheiden der Hufe sowie eine Reihe kleiner Stacheln, die entlang des Schwanzes verliefen. Vermutlich konnten diese Strukturen erhalten bleiben, weil sie sehr rasch mineralisiert wurden. Hierbei könnten auch Aasfresser eine Rolle gespielt haben: Biss- und Fraßspuren an den Mumien, die kurz nach dem Tod entstanden sein müssen, zeigen dreidimensional erhaltene, zusammengesunkene Hautmuster. Durch die Öffnung des Kadavers konnten Verwesungsgase, Flüssigkeiten und Mikroorganismen entweichen, widerstandsfähige Gewebe wie Haut und Krallen dagegen über Wochen oder Monate austrocknen, bevor der Körper endgültig eingebettet und versteinert wurde.117Drumheller, S. K. et al. (2022). Biostratinomic alterations of an Edmontosaurus mummy reveal a pathway for soft tissue preservation without invoking exceptional conditions. PLOS ONE, 17(10), e0275240. doi:10.1371/journal.pone.0275240

 Phillip L. Manning Phillip L. Manning |
In einer Studie aus dem Jahr 2009 berichteten Phillip L. Manning und Kollegen, dass diese Mineralisierung sogar mikroskopisch feine Strukturen wie Zellgrenzen konserviert habe. Außerdem fanden sie chemische Verbindungen mit sogenannten Amidgruppen, die sie als Zerfallsprodukte ehemals vorhandener organischer Moleküle interpretierten. Unversehrte Eiweiße oder innere Organe wie Muskeln, Magen oder andere Eingeweide konnten jedoch nicht nachgewiesen werden.118Manning, P. L. et al. (2009). Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA). Proceedings. Biological Sciences, 276(1672), 3429–3437. doi:10.1098/rspb.2009.0812 |
In einer Folgestudie aus dem Jahr 2015 kamen Manning und sein Team zu der Erkenntnis, dass die Haut von Dinosauriermumien nicht nur als Abdruck erhalten geblieben sei, sondern als mehrere Millimeter dicke Schicht, die noch organisches Material aus dem ursprünglichen Hautgewebe enthalte. Zudem wiesen die Forscher das Farbpigment Melanin nach. Sie betonten jedoch, dass sich daraus noch keine zuverlässige Rekonstruktion der ursprünglichen Körperfärbung ableiten lasse, da viele verschiedene Faktoren die tatsächliche Farbe beeinflussen. Die Verteilung des Melanins könnte künftig aber Hinweise auf ein grobes Hell-Dunkel-Muster des Körpers liefern.119Manning, P. L. et al. (2015). The role and biochemistry of melanin pigment in the exceptional preservation of hadrosaur skin. In Eberth, D. A. & Evans, D. C. (Hgg.), Hadrosaurs. Indiana University Press, Bloomington/Indianapolis, 600–610.
Diese Erkenntnisse habe ich auch in die Farbgebung für „meinen“ Edmontosaurus in Die Weißen Steine einfließen lassen. Er ist damit der einzige Dinosaurier in meiner Geschichte, zu dem wenigstens Indizien vorliegen, wie er wahrscheinlich gefärbt gewesen sein könnte. Allerdings muss ich bewusst „könnte“ schreiben.
 Paul Sereno Paul Sereno |
Paul Sereno und Kollegen kamen in einer Studie aus dem Jahr 2025 zu einem ernüchternderen Ergebnis. Bei zwei neu entdeckten Mumien aus der Lance Formation fanden sie keinerlei Hinweise auf erhaltene organische Verbindungen und Melanin in der Haut. Sie vermuten, dass dies auch für andere berühmte Mumien aus derselben Gesteinseinheit wie „Edmond“ und „Dakota“ gelten muss. Nach ihrer Interpretation besteht die vermeintliche Haut tatsächlich aus einer dünnen Tonschicht, die einen bakteriellen Biofilm überzieht. Dieser Biofilm bildete sich um den Kadaver und konservierte die Oberflächenstruktur der Haut und anderer Weichteile, zwar sehr genau, bevor diese vollständig zerfielen, stellt aber keinen direkten Überrest dar.120Sereno, P. C. et al. (2026). Duck-billed dinosaur fleshy midline and hooves reveal terrestrial clay-template ‘mummification’. Science, 391(6780), eadw3536. doi:10.1126/science.adw3536 |
In einer weiteren Studie aus dem Jahr 2025 gelang bei einem Fossil von Edmontosaurus aber erstmals der eindeutige Nachweis tatsächlich erhaltenen ursprünglichen organischen Materials eines Dinosauriers. Die Forschenden konnten im Fossil Hydroxyprolin nachweisen, eine Aminosäure, die als wichtiger Baustein des Strukturproteins Kollagen dient. Damit widerlegten sie die Hypothese, dass organische Substanzen in Dinosaurierfossilien grundsätzlich auf spätere Verunreinigungen zurückzuführen seien. Stattdessen liefert ihre Untersuchung einen klaren Beleg dafür, dass unter bestimmten Bedingungen originale organische Molekülbestandteile über viele Millionen Jahre erhalten bleiben können.121Tuinstra, L. et al. (2025). Evidence for Endogenous Collagen in Edmontosaurus Fossil Bone. Analytical Chemistry, 97(5), 2618–2628. doi:10.1021/acs.analchem.4c03115
Gehirn
Das Gehirn von Edmontosaurus wurde in mehreren wissenschaftlichen Arbeiten mithilfe sogenannter Endokaste untersucht. Dabei handelt es sich um Nachbildungen des Hohlraums im Schädel, in dem sich einst das Gehirn befand. Solche Untersuchungen wurden sowohl an Edmontosaurus annectens und Edmontosaurus regalis als auch an Exemplaren durchgeführt, die keiner bestimmten Art zugeordnet werden konnten. Die Ergebnisse zeigen, dass das Gehirn für ein so großes Tier vergleichsweise klein war. Der Schädelhohlraum, in dem das Gehirn lag, machte nur etwa ein Viertel der gesamten Schädellänge aus. Je nach Exemplar wurde das Volumen dieses Hohlraums auf 374 bis 450 Milliliter geschätzt. Allerdings füllte das Gehirn den verfügbaren Raum wahrscheinlich nicht vollständig aus. Möglicherweise nahm es nur etwa 50 % des Schädelhohlraums ein, während der restliche Platz von der harten Hirnhaut (Dura mater) und anderen Geweben ausgefüllt wurde. So wird für ein Exemplar mit einem Endokastvolumen von 374 Millilitern ein tatsächliches Gehirnvolumen von lediglich etwa 268 Millilitern angenommen. Das Gehirn selbst war langgestreckt und besaß, wie es für Reptilien typisch ist, keinen Neokortex, also jenen stark entwickelten Bereich der Großhirnrinde, der bei Säugetieren unter anderem für komplexes Denken verantwortlich ist. Ähnlich wie bei Stegosaurus war auch bei Edmontosaurus der Wirbelkanal im Beckenbereich erweitert. Diese Vergrößerung war jedoch deutlich geringer ausgeprägt. Während der sogenannte Endosakralraum bei Stegosaurus etwa 20-mal größer war als der Schädelhohlraum des Gehirns, betrug dieses Verhältnis bei Edmontosaurus lediglich das 2,59-Fache.122Jerison, H. J., Horner, J. R. & Horner, C. C. (2001). Dinosaur forebrains. Journal of Vertebrate Paleontology, 21(3, Suppl.), 64A. doi:10.1080/02724634.2001.10010852
Gliedmaßen und Fortbewegung
Die Gliedmaßenknochen waren länger und schlanker gebaut als bei anderen Hadrosauriern vergleichbarer Größe. Anhand der Proportionen und Form des Schambeins (Pubis) lässt sich zudem ein charakteristisches Becken erkennen.123Chapman, R. E. & Brett-Surman, M. K. (1990). Morphometric observations on hadrosaurid ornithopods. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Perspectives and Approaches. Cambridge University Press, Cambridge, 163–177. Wie andere Hadrosaurier konnte sich Edmontosaurus annectens sowohl zweibeinig als auch vierbeinig fortbewegen. In Die Weißen Steine bin ich der lange verbreiteten Auffassung gefolgt, dass Edmontosaurus sich beim gemächlichen Laufen vierbeinig fortbewegte, doch wenn es die Situation erforderte, zweibeinig die Flucht ergriff.124Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1st ed.). University of California Press, Berkeley, 534–561. Auch Henry Fairfield Osborn nutzte die Skelette im American Museum of Natural History, um beide Fortbewegungsweisen in seinen Rekonstruktionen darzustellen.125Osborn, H. F. (1909). The Upper Cretaceous iguanodont dinosaurs. Nature, 81(2075), 160–162. doi:10.1038/081160a0 Doch heute steht diese Fortbewegungsweise in der Kritik. Der Vergleich von Arm und Beinknochen mit eindeutig zwei oder vierbeinig laufenden Dinosauriern zeigt inzwischen ein klareres Bild: Iguanodon und Ouranosaurus waren zum Beispiel sehr wahrscheinlich dauerhafte Vierbeiner und konnten gar nicht zweibeinig laufen. Bei Lurdusaurus war es sogar noch extremer: Seine kräftigen Ober und Oberschenkelknochen sowie der sehr kurze Mittelfuß deuten auf ein schwerfälliges, säulenartiges Laufen hin und ließen noch nicht einmal zu, dass er sich auch nur temporär auf zwei Beine stellte.126Gônet, J., Allain, R. & Houssaye, A. (2026). Locomotor reassessment of Iguanodon, Ouranosaurus and Lurdusaurus (Dinosauria, Ornithopoda) using osteological correlates. Fossil Record, 29(1), 71–86. doi:10.3897/fr.29.173614 Edmontosaurus war mit diesen früheren Iguanodonten zwar nur entfernt verwandt, teilte jedoch einige ihrer Körpermerkmale. Das zweibeinige Flucht-Laufen ist somit wahrscheinlich eher unwahrscheinlich. Möglicherweise war Edmontosaurus stattdessen zu einer eher Galopp-artigen Gangart fähig.

Wachstum
 David C. Evans David C. Evans
|
In einer Studie aus dem Jahr 2011 erfassten Nicolás Campione und David C. Evans die Daten aller damals bekannten „Edmontosaurus“-Schädel aus dem Campanium und Maastrichtium. Anschließend erstellten sie eine morphometrische Analyse, in der sie verschiedene Schädelmerkmale mit der jeweiligen Schädelgröße verglichen. Ihre Ergebnisse zeigten, dass bei beiden anerkannten Edmontosaurus-Arten viele Merkmale, die zuvor zur Abgrenzung zusätzlicher Arten oder sogar eigener Gattungen verwendet worden waren, direkt mit der Größe des Schädels zusammenhingen. Campione und Evans schlossen daraus, dass sich die Form des Schädels von Edmontosaurus während des Wachstums und der Reifung erheblich veränderte. Heute wissen wir also, dass sich die außergewöhnliche Entenschnabel-Form erst mit dem Erreichen der Geschlechtsreife entwickelte. Mit zunehmendem Alter wurden die Schädel länger und flacher. Nach ihrer Auffassung führten diese altersbedingten Veränderungen in der Vergangenheit zu mehreren Fehlklassifikationen. Die drei zuvor anerkannten Edmontosaurus-Arten aus dem Maastrichtium könnten demnach lediglich verschiedene Entwicklungsstadien einer einzigen Art darstellen: E. saskatchewanensis würde dann die Jungtiere repräsentieren, E. annectens subadulte Tiere und Anatotitan copei vollständig ausgewachsene Individuen.127Campione, N. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186 |

 Mateusz Wosik Mateusz Wosik |
2022 schlugen Mateusz Wosik und David C. Evans auf Grundlage ihrer Untersuchungen verschiedener Exemplare aus unterschiedlichen Fundorten vor, dass Edmontosaurus annectens bereits nach etwa neun Jahren die Geschlechtsreife beziehungsweise das Erwachsenenstadium erreichte. Sie stellten dabei fest, dass dieses Wachstumsmuster gut mit dem anderer Hadrosaurier übereinstimmt.128Wosik, M. & Evans, D. C. (2022). Osteohistological and taphonomic life-history assessment of Edmontosaurus annectens (Ornithischia: Hadrosauridae) from the Late Cretaceous (Maastrichtian) Ruth Mason dinosaur quarry, South Dakota, United States, with implication for ontogenetic segregation between juvenile and adult hadrosaurids. Journal of Anatomy, 241(2), 272–296. doi:10.1111/joa.13679 |
Verhalten
Von Edmontosaurus sind sogenannte Bone Beds bekannt, also Lagerstätten, in denen die Überreste vieler Individuen an einer gemeinsamen Stelle gefunden wurden. Solche Ansammlungen zahlreicher Individuen werden häufig als Hinweis darauf gewertet, dass diese Hadrosaurier gesellig waren und in Gruppen oder Herden lebten.129Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 438–463. Sie stammen sowohl aus der Horseshoe Canyon Formation in Alberta und gehören damit zu Edmontosaurus regalis, aber auch aus der Hell Creek Formation in South Dakota und die Lance Formation in Wyoming, wo Edmontosaurus annectens lebte. Aus letzterer ist ein Bone Bed aus Ton- und Schlammsteinen bekannt, das im Osten Wyomings liegt und sich über eine Fläche von mehr als einem Quadratkilometer erstreckt. Die meisten Knochen konzentrieren sich auf einen etwa 40 Hektar großen Bereich. Dort werden die verstreut liegenden Überreste auf 10.000 bis 25.000 einzelne Edmontosaurus-Individuen geschätzt.130Chadwick, A., Spencer, L. & Turner, L. (2006). Preliminary depositional model for an Upper Cretaceous Edmontosaurus bonebed. Journal of Vertebrate Paleontology, 26(3, Suppl.), 49A. doi:10.1080/02724634.2006.10010069
Die ausgedehnten Vertiefungen rund um die Nasenöffnungen könnten mit sogenannten Nasendivertikeln ausgefüllt gewesen sein. Dabei handelt es sich um aufblasbare Weichteilsäcken, die vermutlich aus der Nasenhöhle hervorgingen. Solche Strukturen könnten sowohl der optischen Kommunikation, etwa durch auffällige Schwellungen, als auch der Lauterzeugung oder Schallverstärkung gedient haben.131Hopson, J. A. (1975). The evolution of cranial display structures in hadrosaurian dinosaurs. Paleobiology, 1(1), 21–43. doi:10.1017/S0094837300002165
Einige Forschende vermuten, dass Edmontosaurus einen Geschlechtsdimorphismus zeigte, also dass Unterschiede zwischen Männchen und Weibchen bestanden. So könnten kräftiger gebaute und schlankere Individuen verschiedene Geschlechter repräsentieren.132Gould, R., Larson, R. & Nellermoe, R. (2003). An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD. Journal of Vertebrate Paleontology, 23(3, Suppl.), 56A–57A. doi:10.1080/02724634.2003.10010538 Bislang konnte dies jedoch nicht eindeutig nachgewiesen werden.
Eine Studie von 2025 beschrieb jedoch auffällige Verletzungsmuster an mittleren Schwanzwirbeln von Hadrosauriern, die wiederholt verheilte Schäden an den Dornfortsätzen zeigen. Eine Finite-Elemente-Analyse eines modellierten Wirbelsäulenabschnitts ergab, dass ein breiter, von oben wirkender Druck die beobachteten Verbiegungen erklären kann. Die verletzte Region entspricht der vermuteten Lage der Kloakenöffnung, was die Möglichkeit nahelegt, dass dieser Druck beim Aufsteigen eines ungestümen Männchens während der Paarung entstand. Solche potenziellen Paarungsverletzungen wären der erste indirekte Hinweis auf Sexualverhalten bei nicht-vogelartigen Dinosauriern und könnten zudem ein neues Mittel bieten, um weibliche Individuen zu erkennen. In Zukunft könnten auch weitere Edmontosaurus-Exemplare mit dieser Methode auf ihr Geschlecht untersucht werden.133Bertozzo, F. et al. (2025). Deciphering causes and behaviors. A recurrent pattern of tail injuries in hadrosaurid dinosaurs. iScience. doi:10.1016/j.isci.2025.113739

 Phil R. Bell Phil R. Bell
|
Aufgrund seiner weiten geografischen Verbreitung, die von Alaska bis Colorado reicht, und weil einige Funde aus der Prince Creek Formation in Regionen lagen, die damals unweit des geografischen Nordpols lagen und in denen während eines großen Teils des Jahres nur wenig Tageslicht herrschte, wird vermutet, dass Edmontosaurus regalis möglicherweise Wanderungen unternahm. In einer Übersichtsarbeit über die Wanderungen von Dinosauriern aus dem Jahr 2008 kamen Phil R. Bell und Eric Snively zu dem Schluss, dass E. regalis zu einer jährlichen Hin- und Rückwanderung von etwa 2.600 Kilometern fähig gewesen sein könnte, sofern er über einen ausreichend hohen Stoffwechsel und genügend Fettreserven verfügte. Eine solche Strecke hätte bei einer Fortbewegungsgeschwindigkeit von ungefähr 2 bis 10 Kilometern pro Stunde zurückgelegt werden können und hätte Wanderungen von Alaska bis nach Alberta ermöglicht. Nach Ansicht von Bell und Snively unterschied sich dieses mögliche Wanderverhalten deutlich von vielen anderen Dinosauriergruppen wie Theropoden, Sauropoden und Ankylosauriern, die ihrer Einschätzung nach den Winter eher in ihren angestammten Lebensräumen verbrachten.134Bell, P. R. & Snively, E. (2008). Polar dinosaurs on parade. A review of dinosaur migration. Alcheringa, 32(3), 271–284. doi:10.1080/03115510802096101 |
 Eric Snively
Eric SnivelyAuch wenn dies für Edmontosaurus annectens noch nicht direkt nachgewiesen ist, ist es dennoch wahrscheinlich, dass auch er ein Herdentier war, und auch für ihn kann ein wanderndes Verhalten durchaus plausibel begründet werden. Es sollte jedoch nicht unerwähnt bleiben, dass es der Wander-Hypothese bereits widersprochen worden ist.
 Anusuya Chinsamy Anusuya Chinsamy |
Anusuya Chinsamy und ihre Kollegen gelangten auf Grundlage von Untersuchungen der Mikrostruktur fossiler Knochen zum Ergebnis, dass die in den Polarregionen lebenden Edmontosaurus-Populationen den Winter wahrscheinlich vor Ort überstanden und nicht regelmäßig in wärmere Gebiete abwanderten.135Chinsamy, A. et al. (2012). Hadrosaurs were perennial polar residents. The Anatomical Record, 295(4), 610–614. doi:10.1002/ar.22428 |
Insofern ist es ebenso möglich, dass vielleicht ein Teil der Population standorttreu lebte, während ein anderer Teil migrierte. Ein ähnliches Verhalten zeigen heute auch manche Zugvögel, bei denen aber nur ein Tei der Population im Winter nach Süden fliegt, wie zum Beispiel Amseln, Stare, Rotkehlchen und Kraniche.
Räuber-Beute-Beziehungen
Tyrannosaurus rex war erwiesenermaßen ein Fressfeind von Edmontosaurus annectens. Ein erwachsenes Exemplar, das heute im Denver Museum of Nature and Science ausgestellt ist, weist Verletzungen am Schwanz auf, die ursprünglich als Spuren eines Angriffs durch einen großen Theropoden gedeutet wurden. Vom Becken aus gezählt zeigen der 13. bis 17. Schwanzwirbel beschädigte Dornfortsätze, die zu einer Attacke von rechts hinten passen könnten. Ein Dornfortsatz wurde teilweise abgeschert, andere sind verbogen, und an drei Stellen befinden sich Vertiefungen, die als mögliche Zahnabdrücke interpretiert wurden. Da der Schwanz an dieser Stelle mindestens 2,9 Meter hoch gewesen wäre und aus derselben Gesteinsformation kein anderer ausreichend großer Raubsaurier bekannt ist, würde nur Tyrannosaurus rex als Verursacher infrage kommen. Die Knochen zeigen Anzeichen einer teilweisen Heilung, allerdings starb das Tier, bevor die Verletzungen vollständig verheilen konnten. Außerdem finden sich Hinweise auf eine Knocheninfektion. Kenneth Carpenter, der das Exemplar untersuchte, bemerkte zusätzlich einen bereits deutlich besser verheilten Bruch an der linken Hüfte. Er vermutete deshalb, dass das Tier möglicherweise hinkte und dadurch zu einem leichteren Ziel wurde. Da Edmontosaurus den Vorfall offenbar zunächst überlebte, schlug Carpenter vor, dass er seinem Angreifer entkommen oder ihn sogar im Kampf abgewehrt haben könnte.136Carpenter, K. (1998). Evidence of predatory behavior by theropod dinosaurs. Gaia, 15, 135–144. Allerdings kommen auch andere Ursachen als der Angriff eines Beutegreifers für diese Verletzungen in Betracht, wie etwa ein Unfall oder ein Gerangel mit einem Artgenossen.137Siviero, A. R. T. V. et al. (2020). Skeletal trauma with implications for intratail mobility in Edmontosaurus annectens from a monodominant bonebed, Lance Formation (Maastrichtian), Wyoming, USA. PALAIOS, 35(4), 201–214. doi:10.2110/palo.2019.079

Etwas eindeutiger sind Verletzungsspuren an einem weiteren, noch jugendlichen Exemplar aus South Dakota, das etwa 7,6 Meter lang war, als es starb. Es weist Zahnspuren kleinerer Theropoden am Unterkiefer auf, ein Teil dieser Verletzungen war bereits verheilt. Michael Triebold vermutete, dass kleinere Raubsaurier den Halsbereich des Tieres attackiert hatten. Seiner Interpretation zufolge überlebte Edmontosaurus den ersten Angriff zunächst, erlag aber kurze Zeit später seinen Verletzungen.138Campagna, T. (2000). The PT interview. Michael Triebold. Prehistoric Times, 40, 18–19. Eine 2026 veröffentlichte Studie beschrieb außerdem ein Exemplar von Edmontosaurus annectens, bei dem ein Zahn eines Tyrannosauriers noch im Nasenbein steckte. Die Forschenden interpretierten diesen Fund als wahrscheinliches Ergebnis eines Bisses in die Schnauze während eines Raubversuchs kurz vor dem Tod des Tieres.139Wyenberg-Henzler, T. C. A. & Scannella, J. B. (2026). Behavioral implications of an embedded tyrannosaurid tooth and associated tooth marks on an articulated skull of Edmontosaurus from the Hell Creek Formation, Montana. PeerJ, 14, e20796. doi:10.7717/peerj.20796

Quellenangaben:
- 1Bakker, R. T. (1986). The Dinosaur Heresies. 438.
- 2Lehman, T. M. (2001). Late Cretaceous dinosaur provinciality. In Tanke, D. & Carpenter, K. (Hgg.), Mesozoic Vertebrate Life. Indiana University Press, Bloomington/Indianapolis, 310–328.
- 3Weishampel, D. B. et al. (2004). Dinosaur distribution. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 517–606.
- 4Russell, D. A. (1989). An Odyssey in Time. Dinosaurs of North America. 175–180.
- 5Osborn, H. F. (1909). The Upper Cretaceous iguanodont dinosaurs. Nature, 81(2075), 160–162. doi:10.1038/081160a0
- 6Norell, M. A., Gaffney, E. S. & Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. Knopf, New York, 156–158.
- 7Glut, D. F. (1997). Anatotitan. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 132–134.
- 8Cope, E. D. (1883). On the characters of the skull in the Hadrosauridae. Proceedings of the Philadelphia Academy of Natural Sciences, 35, 97–107.
- 9Leidy, J. (1856). Notice of remains of extinct reptiles and fishes, discovered by F. V. Hayden in the Bad Lands of the Judith River, Nebraska Territories. Proceedings of the Academy of Natural Science Philadelphia, 8, 72–73.
- 10Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.
- 11Marsh, O. C. (1892). Restorations of Claosaurus and Ceratosaurus. American Journal of Science, 44(262), 343–349. doi:10.2475/ajs.s3-44.262.343
- 12Marsh, O. C. (1890). Additional characteristics of the Ceratopsidae, with notice of new Cretaceous dinosaurs. American Journal of Science, 3rd Series, 39(233), 418–426. doi:10.2475/ajs.s3-39.233.418
- 13Marsh, O. C. (1892). Notice of new reptiles from the Laramie Formation. American Journal of Science, 43(257), 449–453. doi:10.2475/ajs.s3-43.257.449
- 14Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.
- 15Marsh, O. C. (1892). Restorations of Claosaurus and Ceratosaurus. American Journal of Science, 44(262), 343–349. doi:10.2475/ajs.s3-44.262.343
- 16Lucas, F. A. (1904). The dinosaur Trachodon annectens. Smithsonian Miscellaneous Collections, 45, 317–320.
- 17Norell, M. A., Gaffney, E. S. & Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. Knopf, New York, 154–155.
- 18Dal Sasso, C. & Brillante, G. (2004). Dinosaurs of Italy. Indiana University Press, Bloomington and Indianapolis, 112.
- 19Norell, M. A., Gaffney, E. S. & Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. Knopf, New York, 156–158.
- 20Hatcher, J. B. (1902). The genera and species of the Trachodontidae (Hadrosauridae, Claosauridae) Marsh. Annals of the Carnegie Museum, 1(14), 377–386. doi:10.5962/p.331063
- 21Gilmore, C. W. (1915). On the genus Trachodon. Science, 41(1061), 658–660. doi:10.1126/science.41.1061.658
- 22Leidy, J. (1856). Notice of extinct Vertebrata, discovered by F. V. Hayden during the expedition to the Sioux country under the command of Lieut. G. K. Warren. Proceedings of the Academy of Natural Science Philadelphia, 8, 311–312.
- 23Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.
- 24Cope, E. D. (1875). Report on the vertebrate paleontology of Colorado. US Government Printing Office.
- 25Leidy, J. (1858). Hadrosaurus foulkii, a new saurian from the Cretaceous of New Jersey, related to Iguanodon. Proceedings of the Academy of Natural Sciences of Philadelphia, 10, 213–218.
- 26Lucas, F. A. (1900). Paleontological Notes. Science, 12(308), 809–810.
- 27Gilmore, C. W. (1915). On the genus Trachodon. Science, 41(1061), 658–660. doi:10.1126/science.41.1061.658
- 28Gilmore, C. W. (1924). A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta. Canada Department of Mines Geological Survey Bulletin (Geological Series), 38(43), 13–26.
- 29Russell, L. S. (1930). Upper Cretaceous dinosaur faunas of North America. Proceedings of the American Philosophical Society, 69(1), 133–159.
- 30Sternberg, C. M. (1936). The systematic position of Trachodon. Journal of Paleontology, 10(7), 652–655.
- 31Sternberg, C. M. (1939). Were there Proboscis-bearing Dinosaurs? Discussion of Cranial Protuberances in the Hadrosauridae. Journal of Natural History, 3(17), 556–560.
- 32Marsh, O. C. (1897). Vertebrate fossils of the Denver Basin. U.S. Geological Survey, Monthly, 27, 473–527.
- 33Sternberg, C. M. (1926). A new species of Thespesius from the Lance Formation of Saskatchewan. Canada Department of Mines Geological Survey Bulletin (Geological Series), 44(46), 73–84.
- 34Lambe, L. M. (1917). A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta. The Ottawa Naturalist, 31(7), 65–73.
- 35Lambe, L. M. (1902). On Vertebrata of the mid-Cretaceous of the Northwest Territory. New genera and species from the Belly River Series (mid-Cretaceous). Contributions to Canadian Paleontology, 3, 25–81.
- 36Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272.
- 37Lambe, L. M. (1920). The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta. Memoir, 120. Department of Mines, Geological Survey of Canada, 1–79.
- 38Cope, E. D. (1871). Supplement to the synopsis of the extinct Batrachia and Reptilia of North America. Proceedings of the American Philosophical Society, 12(86), 41–52.
- 39Glut, D. F. (1997). Edmontosaurus. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 389–396.
- 40Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 438–463.
- 41Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1. Aufl.). University of California Press, Berkeley, 534–561.
- 42Cope, E. D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. U.S. Geological and Geographical Survey of the Territories Annual Report, 1, 9–28.
- 43Cope, E. D. (1874). Report on the vertebrate paleontology of Colorado. U.S. Geological and Geographical Survey of the Territories Annual Report, 2, 429–454.
- 44Carpenter, K. & Young, D. B. (2002). Late Cretaceous dinosaurs from the Denver Basin, Colorado. Rocky Mountain Geology, 37(2), 237–254. doi:10.2113/11
- 45Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 438–463.
- 46Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1. Aufl.). University of California Press, Berkeley, 534–561.
- 47Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272.
- 48Glut, D. F. (1982). The New Dinosaur Dictionary. Citadel Press, Secaucus, NJ, 57.
- 49Brett-Surman, M. K. (1975). The appendicular anatomy of hadrosaurian dinosaurs. M.A. thesis, University of California, Berkeley.
- 50Brett-Surman, M. K. (1979). Phylogeny and paleobiogeography of hadrosaurian dinosaurs. Nature, 277(5697), 560–562. doi:10.1038/277560a0
- 51Brett-Surman, M. K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation, George Washington University, Washington, D.C.
- 52Glut, D. F. (1982). The New Dinosaur Dictionary. Citadel Press, Secaucus, NJ, 49, 53.
- 53Lambert, D. & the Diagram Group (1983). A Field Guide to Dinosaurs. Avon Books, New York, 156–161.
- 54Chapman, R. E. & Brett-Surman, M. K. (1990). Morphometric observations on hadrosaurid ornithopods. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Perspectives and Approaches. Cambridge University Press, Cambridge, 163–177.
- 55Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1st ed.). University of California Press, Berkeley, 534–561.
- 56Olshevsky, G. (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. Mesozoic Meanderings, 2. Publications Requiring Research, San Diego.
- 57Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2nd ed.). University of California Press, Berkeley, 438–463.
- 58Lee, C. (2007). Scientists Get Rare Look at Dinosaur Soft Tissue. The Washington Post.
- 59Campione, N. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186
- 60Campione, N. S. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186
- 61Glut, D. F. (1997). Anatotitan. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 132–134.
- 62Campione, N. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186
- 63Brett-Surman, M. K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation, George Washington University, Washington, D.C.
- 64Mori, H., Druckenmiller, P. S. & Erickson, G. M. (2015). A new Arctic hadrosaurid from the Prince Creek Formation (lower Maastrichtian) of northern Alaska. Acta Palaeontologica Polonica, 61. doi:10.4202/app.00152.2015
- 65Xing, H., Mallon, J. C. & Currie, M. L. (2017). Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLOS ONE, 12(4), e0175253. doi:10.1371/journal.pone.0175253
- 66Takasaki, R. et al. (2020). Re-examination of the cranial osteology of the Arctic Alaskan hadrosaurine with implications for its taxonomic status. PLOS ONE, 15(5), e0232410. doi:10.1371/journal.pone.0232410
- 67Sharpe, H. S. et al. (2023). Craniomandibular anatomy of a juvenile specimen of Edmontosaurus regalis Lambe, 1917 clarifies issues in ontogeny and biogeography. Journal of Vertebrate Paleontology, 43(5), e2326644. doi:10.1080/02724634.2024.2326644
- 68Davies, K. L. (1983). Hadrosaurian dinosaurs of Big Bend National Park. M.A. thesis, University of Texas, Austin, 231 S.
- 69Weishampel, D. B. et al. (2004). Dinosaur distribution. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 517–606.
- 70Wagner, J. R. (2001). The hadrosaurian dinosaurs (Ornithischia: Hadrosauria) of Big Bend National Park, Brewster County, Texas, with implications for Late Cretaceous paleozoogeography. M.Sc. thesis, Texas Tech University, Austin, Texas.
- 71Lehman, T. M., Wick, S. L. & Wagner, J. R. (2016). Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation, Big Bend National Park, Texas. Journal of Paleontology, 90(2), 1–24. doi:10.1017/jpa.2016.48
- 72Cope, E. D. (1969). Synopsis of the extinct batrachia, reptilia and aves of North America. Transactions of the American Philosophical Society, 14.
- 73Parks, W. A. (1923). Corythosaurus intermedius, a new species of trachodont dinosaur. University of Toronto Geological Studies, 15, 5–57.
- 74Prieto-Márquez, A. (2010). Global phylogeny of Hadrosauridae (Dinosauria. Ornithopoda) using parsimony and Bayesian methods. Zoological Journal of the Linnean Society, 159(2), 435–502. doi:10.1111/j.1096-3642.2009.00617.x
- 75Pantelides, A., Upchurch, P., Mannion, P. D., Gravanis, E., Henderson, D. M. & Kyriakidis, P. (2026). ENHYDROSS – a new mechanistic model supports the trans-oceanic dispersal capability of terrestrial vertebrates. Ecology and Evolution, 16, e73280. doi:10.1002/ece3.73280
- 76Cope, E. D. (1883). On the characters of the skull in the Hadrosauridae. Proceedings of the Philadelphia Academy of Natural Sciences, 35, 97–107.
- 77Lull, R. S. & Wright, N. E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper, 40. Geological Society of America, 225.
- 78Osborn, H. F. (1909). The Upper Cretaceous iguanodont dinosaurs. Nature, 81(2075), 160–162. doi:10.1038/081160a0
- 79Morris, W. J. (1970). Hadrosaurian dinosaur bills. Morphology and function. Contributions in Science (Los Angeles County Museum of Natural History), 193, 1–14.
- 80Sues, H.-D. (1997). Ornithopods. In Farlow, J. O. & Brett-Surman, M. K. (Hgg.), The Complete Dinosaur. Indiana University Press, Bloomington, 338.
- 81Glut, D. F. (1997). Anatotitan. In Dinosaurs: The Encyclopedia. McFarland & Co., Jefferson, North Carolina, 132–134.
- 82Horner, J. R., Goodwin, M. B. & Myhrvold, N. (2011). Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLOS ONE, 6(2), e16574. doi:10.1371/journal.pone.0016574
- 83Prieto-Márquez, A. (2014). A juvenile Edmontosaurus from the late Maastrichtian (Cretaceous) of North America. Implications for ontogeny and phylogenetic inference in saurolophine dinosaurs. Cretaceous Research, 50, 282–303. doi:10.1016/j.cretres.2014.05.003
- 84Jiménez-Moreno, F. J. et al. (2025). First Population Analysis in Hadrosauroid dinosaurs (Ornithopoda: Iguanodontia: Hadrosauroidea). Evolving Earth, 3, 100072. doi:10.1016/j.eve.2025.100072
- 85Wosik, M. et al. (2022). Osteohistological and taphonomic life-history assessment of Edmontosaurus annectens (Ornithischia: Hadrosauridae) from the Late Cretaceous (Maastrichtian) Ruth Mason dinosaur quarry, South Dakota, United States, with implication for ontogenetic segregation between juvenile and adult hadrosaurids. Journal of Anatomy, 241(2), 272–296. doi:10.1111/joa.13679
- 86Chapman, R. E. & Brett-Surman, M. K. (1990). Morphometric observations on hadrosaurid ornithopods. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Perspectives and Approaches. Cambridge University Press, Cambridge, 163–177.
- 87Cope, E. D. (1883). On the characters of the skull in the Hadrosauridae. Proceedings of the Philadelphia Academy of Natural Sciences, 35, 97–107.
- 88Morris, W. J. (1970). Hadrosaurian dinosaur bills. Morphology and function. Contributions in Science (Los Angeles County Museum of Natural History), 193, 1–14.
- 89Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272.
- 90Ostrom, J. H. (1964). A reconsideration of the paleoecology of the hadrosaurian dinosaurs. American Journal of Science, 262(8), 975–997. doi:10.2475/ajs.262.8.975
- 91Barrett, P. M. (2005). The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria). Palaeontology, 48(2), 347–358. doi:10.1111/j.1475-4983.2005.00448.x
- 92Williams, V. S., Barrett, P. M. & Purnell, M. A. (2009). Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. Proceedings of the National Academy of Sciences, 106(27), 11194–11199. doi:10.1073/pnas.0812631106
- 93Kubo, T. et al. (2023). Dental microwear texture analysis reveals a likely dietary shift within Late Cretaceous ornithopod dinosaurs. Palaeontology, 66(6), e12681. doi:10.1111/pala.12681
- 94Fiorillo, A. R. (2011). Microwear patterns on the teeth of northern high latitude hadrosaurs with comments on microwear patterns in hadrosaurs as a function of latitude and seasonal ecological constraints. Palaeontologia Electronica, 14(3), 1–17.
- 95Erickson, G. M. et al. (2012). Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science, 338(6103), 98–101. doi:10.1126/science.1224495
- 96Rybczynski, N. et al. (2008). A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses. Palaeontologia Electronica, 11(2), online publication.
- 97Williams, V. S., Barrett, P. M. & Purnell, M. A. (2009). Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. Proceedings of the National Academy of Sciences, 106(27), 11194–11199. doi:10.1073/pnas.0812631106
- 98Holliday, C. M. & Witmer, L. M. (2008). Cranial kinesis in dinosaurs. Intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids. Journal of Vertebrate Paleontology, 28(4), 1073–1088. doi:10.1671/0272-4634-28.4.1073
- 99Rybczynski, N. et al. (2008). A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses. Palaeontologia Electronica, 11(2), online publication.
- 100Williams, V. S., Barrett, P. M. & Purnell, M. A. (2009). Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. Proceedings of the National Academy of Sciences, 106(27), 11194–11199. doi:10.1073/pnas.0812631106
- 101Cuthbertson, R. S. et al. (2012). Kinetic limitations of intracranial joints in Brachylophosaurus canadensis and Edmontosaurus regalis (Dinosauria: Hadrosauridae), and their implications for the chewing mechanics of hadrosaurids. The Anatomical Record, 295(6), 968–979. doi:10.1002/ar.22458
- 102Creisler, B. S. (2007). Deciphering duckbills. A history in nomenclature. In Carpenter, K. (Hg.), Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, 185–210.
- 103Naish, D. (2018). Enough with the Duck-Billed Dinosaurs. Scientific American.
- 104Black, R. (2012). Shovel-Beaked, Not Duck-Billed. Smithsonian Magazine.
- 105Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2nd ed.). University of California Press, Berkeley, 438–463.
- 106Sternberg, C. H. (1909). A new Trachodon from the Laramie Beds of Converse County, Wyoming. Science, 29(749), 753–754. doi:10.1126/science.29.749.747
- 107Kräusel, R. (1922). Die Nahrung von Trachodon. Paläontologische Zeitschrift, 4(2–3), 80. doi:10.1007/BF03041547
- 108Abel, O. (1922). Diskussion zu den Vorträgen R. Kräusel and F. Versluys. Paläontologische Zeitschrift, 4(2–3), 87. doi:10.1007/BF03041547
- 109Wieland, G. R. (1925). Dinosaur feed. Science, 61(1589), 601–603. doi:10.1126/science.61.1589.601
- 110Ostrom, J. H. (1964). A reconsideration of the paleoecology of the hadrosaurian dinosaurs. American Journal of Science, 262(8), 975–997. doi:10.2475/ajs.262.8.975
- 111Carpenter, K. (1987). Paleoecological significance of droughts during the Late Cretaceous of the Western Interior. In Currie, P. J. & Koster, E. H. (Hgg.), Fourth Symposium on Mesozoic Terrestrial Ecosystems, Drumheller, August 10–14, 1987 (Occasional Paper of the Tyrrell Museum of Palaeontology 3). Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, 42–47.
- 112Carpenter, K. (2007). How to make a fossil. Part 2. Dinosaur mummies and other soft tissue. The Journal of Paleontological Sciences, online.
- 113Currie, P. J., Koppelhus, E. B. & Muhammad, A. F. (1995). Stomach contents of a hadrosaurid from the Dinosaur Park Formation (Campanian, Upper Cretaceous) of Alberta, Canada. In Sun, A. & Wang, Y. (Hgg.), Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. China Ocean Press, Beijing, 111–114.
- 114Tweet, J. S. et al. (2008). Probable gut contents within a specimen of Brachylophosaurus canadensis (Dinosauria: Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana. PALAIOS, 23(9), 624–635. doi:10.2110/palo.2007.p07-044r
- 115Stanton Thomas, K. J. & Carlson, S. J. (2004). Microscale δ¹⁸O and δ¹³C isotopic analysis of an ontogenetic series of the hadrosaurid dinosaur Edmontosaurus. Implications for physiology and ecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 206, 257–287. doi:10.1016/j.palaeo.2004.01.007
- 116Lull, R.S. & Wright, N.E. (1942). „Hadrosaurian dinosaurs of North America.“ Geological Society of America Special Papers, 40: 1-272.
- 117Drumheller, S. K. et al. (2022). Biostratinomic alterations of an Edmontosaurus mummy reveal a pathway for soft tissue preservation without invoking exceptional conditions. PLOS ONE, 17(10), e0275240. doi:10.1371/journal.pone.0275240
- 118Manning, P. L. et al. (2009). Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA). Proceedings. Biological Sciences, 276(1672), 3429–3437. doi:10.1098/rspb.2009.0812
- 119Manning, P. L. et al. (2015). The role and biochemistry of melanin pigment in the exceptional preservation of hadrosaur skin. In Eberth, D. A. & Evans, D. C. (Hgg.), Hadrosaurs. Indiana University Press, Bloomington/Indianapolis, 600–610.
- 120Sereno, P. C. et al. (2026). Duck-billed dinosaur fleshy midline and hooves reveal terrestrial clay-template ‘mummification’. Science, 391(6780), eadw3536. doi:10.1126/science.adw3536
- 121Tuinstra, L. et al. (2025). Evidence for Endogenous Collagen in Edmontosaurus Fossil Bone. Analytical Chemistry, 97(5), 2618–2628. doi:10.1021/acs.analchem.4c03115
- 122Jerison, H. J., Horner, J. R. & Horner, C. C. (2001). Dinosaur forebrains. Journal of Vertebrate Paleontology, 21(3, Suppl.), 64A. doi:10.1080/02724634.2001.10010852
- 123Chapman, R. E. & Brett-Surman, M. K. (1990). Morphometric observations on hadrosaurid ornithopods. In Carpenter, K. & Currie, P. J. (Hgg.), Dinosaur Systematics. Perspectives and Approaches. Cambridge University Press, Cambridge, 163–177.
- 124Weishampel, D. B. & Horner, J. R. (1990). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (1st ed.). University of California Press, Berkeley, 534–561.
- 125Osborn, H. F. (1909). The Upper Cretaceous iguanodont dinosaurs. Nature, 81(2075), 160–162. doi:10.1038/081160a0
- 126Gônet, J., Allain, R. & Houssaye, A. (2026). Locomotor reassessment of Iguanodon, Ouranosaurus and Lurdusaurus (Dinosauria, Ornithopoda) using osteological correlates. Fossil Record, 29(1), 71–86. doi:10.3897/fr.29.173614
- 127Campione, N. E. & Evans, D. C. (2011). Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae). Implications for Latest Cretaceous Megaherbivore Diversity in North America. PLOS ONE, 6(9), e25186. doi:10.1371/journal.pone.0025186
- 128Wosik, M. & Evans, D. C. (2022). Osteohistological and taphonomic life-history assessment of Edmontosaurus annectens (Ornithischia: Hadrosauridae) from the Late Cretaceous (Maastrichtian) Ruth Mason dinosaur quarry, South Dakota, United States, with implication for ontogenetic segregation between juvenile and adult hadrosaurids. Journal of Anatomy, 241(2), 272–296. doi:10.1111/joa.13679
- 129Horner, J. R., Weishampel, D. B. & Forster, C. A. (2004). Hadrosauridae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hgg.), The Dinosauria (2. Aufl.). University of California Press, Berkeley, 438–463.
- 130Chadwick, A., Spencer, L. & Turner, L. (2006). Preliminary depositional model for an Upper Cretaceous Edmontosaurus bonebed. Journal of Vertebrate Paleontology, 26(3, Suppl.), 49A. doi:10.1080/02724634.2006.10010069
- 131Hopson, J. A. (1975). The evolution of cranial display structures in hadrosaurian dinosaurs. Paleobiology, 1(1), 21–43. doi:10.1017/S0094837300002165
- 132Gould, R., Larson, R. & Nellermoe, R. (2003). An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD. Journal of Vertebrate Paleontology, 23(3, Suppl.), 56A–57A. doi:10.1080/02724634.2003.10010538
- 133Bertozzo, F. et al. (2025). Deciphering causes and behaviors. A recurrent pattern of tail injuries in hadrosaurid dinosaurs. iScience. doi:10.1016/j.isci.2025.113739
- 134Bell, P. R. & Snively, E. (2008). Polar dinosaurs on parade. A review of dinosaur migration. Alcheringa, 32(3), 271–284. doi:10.1080/03115510802096101
- 135Chinsamy, A. et al. (2012). Hadrosaurs were perennial polar residents. The Anatomical Record, 295(4), 610–614. doi:10.1002/ar.22428
- 136Carpenter, K. (1998). Evidence of predatory behavior by theropod dinosaurs. Gaia, 15, 135–144.
- 137Siviero, A. R. T. V. et al. (2020). Skeletal trauma with implications for intratail mobility in Edmontosaurus annectens from a monodominant bonebed, Lance Formation (Maastrichtian), Wyoming, USA. PALAIOS, 35(4), 201–214. doi:10.2110/palo.2019.079
- 138Campagna, T. (2000). The PT interview. Michael Triebold. Prehistoric Times, 40, 18–19.
- 139Wyenberg-Henzler, T. C. A. & Scannella, J. B. (2026). Behavioral implications of an embedded tyrannosaurid tooth and associated tooth marks on an articulated skull of Edmontosaurus from the Hell Creek Formation, Montana. PeerJ, 14, e20796. doi:10.7717/peerj.20796
Hat dir dieser Beitrag gefallen?
Für noch mehr Lesestoff schau gerne in die Kategorie Meine Bücher. Alle meine Werke findest du auch in der Sidebar rechts. Dort gibt es auch einen Direktlink zu Amazon, wo du sie gleich bestellen kannst. ➡️
Wie kannst du Die Weißen Steine sonst noch unterstützen?
Klar, in erster Linie natürlich, wenn du dir eines meiner Bücher kaufst. Aber du kannst mir auch auf anderen Wegen eine große Freude machen:
- Teile diesen Beitrag auf deinen Social-Media-Kanälen!
- Schreibe mir eine nette Rezension auf Amazon!
- Kaufe hier auf der Website über Affiliate Links ein!
Ich kriege übrigens auch eine kleine Unterstützung, wenn du etwas völlig anderes kaufst. Es muss nicht unbedingt der hier beworbene Artikel sein! Jeder kleine Centbetrag hilft, diese Seite am Laufen zu erhalten.
 |
Zusammen mit meiner lieben Frau Sahar habe ich auch noch eine Menge mehr zu bieten! Sahar ist nämlich eine großartige Künstlerin und fertigt Gemälde, Lesezeichen, Stofftaschen und viele andere coole Artikel an. Auch mit tollen Urzeit-Motiven! Klick dich einfach mal durch die Beiträge in der Kategorie Produkte & Services. Bestimmt findest du da etwas nach deinem Geschmack. Oder auch ein schönes Geschenk für deine Lieben. |
Einen Grund, anderen eine Freude zu bereiten, findet man schließlich immer!
Spannend und informativ zu lesen. Bin gespannt auf den zweiten Band
Vielen Dank! Band II kommt vorraussichtlich im Juli.