
Name: Triceratops prorsus („Schlichtes Dreihorngesicht“)
Beschrieben: 1890 von Othniel C. Marsh
Ordnung: Ornithischia; Familie: Ceratopsidae
Länge: ♂ bis zu 8,3, ♀ bis 9m
Gewicht: ♂ bis zu 8.000kg, ♀ bis zu 9.600kg
Ernährung: herbivor
Beschreibung:
Triceratops ist einer der berühmtesten Dinosaurier und bei weitem der häufigste in ganz Laramidia. Während der Trockenzeit sammeln sich viele Tiere im mittleren Abschnitt des westlichen amerikanischen Kontinents, wo es zu dieser Jahreszeit noch warm, aber auch noch deutlich grüner ist als in ihren übrigen Verbreitungsgebieten. Von dort aus ziehen sie dann gemeinsam in riesigen Herdenverbänden weiter südlich, wo sie dann den Winter verbringen. Im Sommer trifft man ihn auch wieder in den nördlichen Breiten Laramidias an, wo er sich in weitläufigen Prärien und lichten Wäldern aufhält. Seine Lieblingsspeise sind niedrig wachsende Pflanzen, Sträucher und auch Wurzelknollen, welche die Tiere mit ihren starken Hornschnäbeln aus dem Boden wühlen. In seltenen Fällen sieht man sie allerdings auch an Kadavern anderer verendeter Großdinosaurier. Fleisch gehört aber wirklich nur in Ausnahmefällen auf ihre Speisekarte.

Triceratops-Männchen sind braunrot gefärbt mit dunkelroten Streifen. Ihr beeindruckender Nackenschild ist mit Keratin und schillernden Farbmustern verziert. Die Weibchen sind weniger farbenfroh. Ihre Haut ist graubraun, bei manchen Individuen beinahe schwarz. Während der fruchtbaren Sommermonate bilden die Weibchen kleinere Verbände, die sich über den ganzen Kontinent verteilen. Die Herden umfassen mehrere weibliche Individuen, die eng miteinander verwandt sind. Gemeinsam mit ihren Müttern, Schwestern und Tanten sowie ihrem mindestens eineinhalbjährigen Nachwuchs legen sie auf ihren Wanderungen jedes Jahr tausende von Kilometern zurück. Nur die Mütter, die im Frühjahr frischen Nachwuchs bekommen haben, lassen diese Wanderungen aus. Männchen ziehen meist als Einzelgänger oder in kleinen Gruppen durch ihr Revier.

Lebensweise:
Im Frühjahr schlüpfen aus den ledrigen, spitz zulaufenden Eiern, die ein Triceratops-Weibchen in ihrem Nisthügel bewacht, bis zu 15 Jungtiere. Die Mütter verbleiben mit ihnen bis zum übernächsten Herbst in den Brutgebieten. Erst dann sind die Kleinen nämlich kräftig genug, um den langen Marsch zu wagen. Trotz der hohen Nachkommenzahl erreichen nur wenige Jungtiere das Erwachsenenalter. Die Mütter kümmern sich liebevoll um ihren Nachwuchs, und das bis zu fünf Jahre lang, in denen ein Junge führendes Weibchen auch keine neuen Eier legt. Während dieser Zeit verteidigt sie ihre Jungtiere vehement gegen Fressfeinde und duldet keine fremden Männchen in der Nähe des Nachwuchses. Zur Verteidigung gegen Raubtiere bilden die Weibchen einen schützenden Ring um ihre Jungen. Dabei stellen die Tiere ihre imposanten Hörner und Nackenschilde zur Schau, eine Drohgebärde, die in den meisten Fällen Wirkung zeigt.

Die Männchen legen ein deutlich einsameres Leben an den Tag: Meist ziehen bloß ein bis zwei jüngere Männchen mit einem weiteren, deutlich älteren Veteranen umher, um sich gegenseitig vor Feinden zu schützen. In der Paarungszeit werden aus den Freunden jedoch stets erbitterte Rivalen! Dann werden die Männchen nämlich extrem aggressiv und greifen alles an, was ihnen bedrohlich erscheint. Und das kann im Grunde jedes andere Tier sein, selbst so mancher Baumstumpf hat bereits den Zorn eines wütenden Triceratops zu spüren bekommen. Bei den Brunftkämpfen ringen die Männchen erbittert miteinander um die Herden der Weibchen, indem sie ihre Hörner verhaken und solange schieben, bis einer der Kontrahenten nachgibt. Diese kräftezehrenden Kämpfe enden zwar selten tödlich, viele Männchen sterben jedoch trotzdem, an Entkräftung oder weil sie geschwächt zur leichten Beute für Räuber werden. Die Überlebenden kehren nach der Brunft in ihre Reviere zurück.

Triceratops in Die Weißen Steine:
 |
Band I:Triceratops tritt im Kapitel „Schräge Vögel“ in Erscheinung. Eine riesige Herde weiblicher Tiere, die sich bereits zu ihrer Herbstwanderung sammelt, wird dort von den Jungs aus der Ferne gesichtet und für eine Rinderherde gehalten. |
 |
Band II:Eine ganze Herde Triceratops begegnet Marie, Leon und Heinrich im Kapitel „Die Wesen am Fluss“ und überzeugt Heinrich schließlich, in der Dinosaurierzeit gelandet zu sein. Die gleiche Herde verwüstet mehrmals das Lager der anderen Schüler, die sich am Seeufer niedergelassen haben, und zwingt sie im Kapitel „Elementare Dinge“ schließlich dazu, sich einen neuen Lagerplatz zu suchen. Im Kapitel „Verbotene Früchte“ freundet sich Hannah schließlich mit einem Jungtier an, das sie liebevoll auf den Namen „Trisha“ tauft. |
 |
Band III:Moritz kann Triceratops oft in der Nähe des Sees beobachten und macht dabei viele interessante Entdeckungen über die Tiere, die er in seinem Tagebuch – einst Alinas Taschenkalender – festhält. Melina fertigt dazu später Zeichnungen an. Eine der gezeichneten Begegnungen hat es in sich: Im Kapitel „Der gehörnte Koloss“ greift ein junger Triceratops-Bulle Moritz und Ramzi bei der Nahrungssuche an. Moritz kann ihm nur mit knapper Not und durch seine Schwimmkünste entkommen. |
 |
Band IV:Hannah trifft in „Erinnerungen“ Trisha wieder, die seit ihrer letzten Begegnung deutlich gewachsen ist, ihre menschliche Freundin aber trotzdem noch wiedererkennt. Später werden Leon, Marie, Wasi, Hannah und John Zeuge einer dramatischen Jagdszene. Die T. rex-Familie erlegt dabei gemeinsam einen jungen, männlichen Triceratops. In „Der sterbende See“ befinden sich auch einige Triceratops unter den fliehenden Tieren, vor denen Moritz und seine Freunde in Deckung gehen müssen. Marie und ihre Freunde sichten die Herde in „Heimkehr“ ein weiteres Mal, als sie mit dem bewusstlosen Max über die Ebene wandern. |
 |
Hier geht’s zurück zur Hauptseite Leben in der Urzeit! |
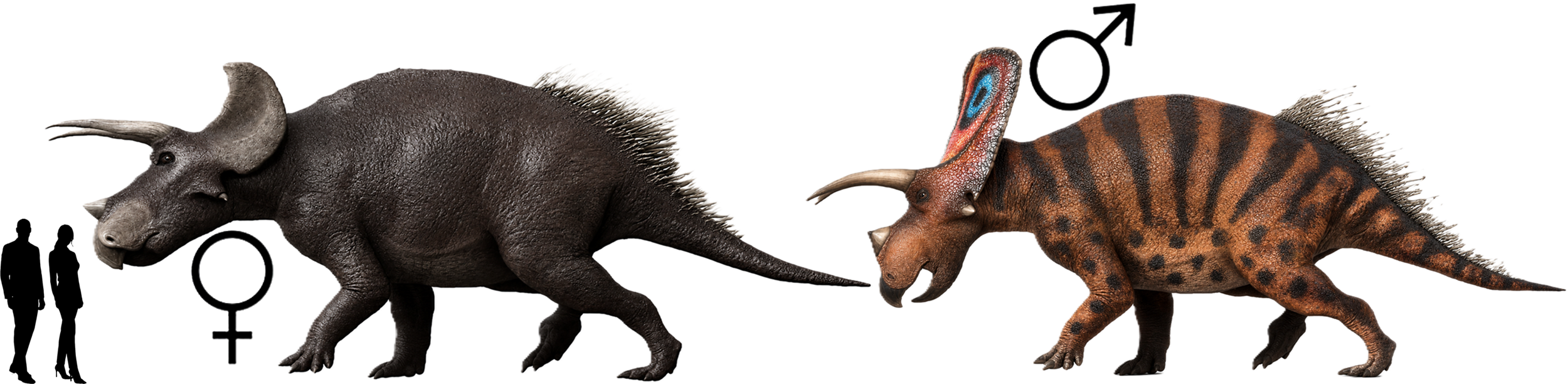
Wissenschaftliches zu Triceratops:
 |
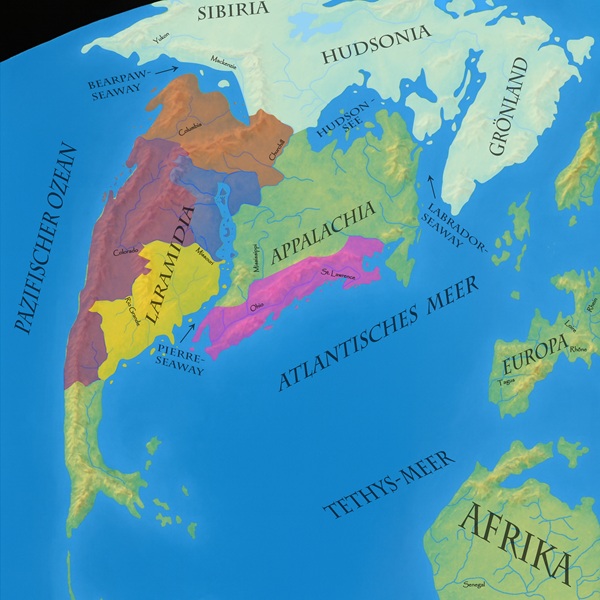
Triceratops ist eines der am längsten bekannten Lebewesen des kreidezeitlichen Nordamerikas und einer der am häufigsten dort gefundenen Dinosaurier. Seine Fossilien wurden in vielen Lagerstätten des westlichen Nordamerika, das wir zu dieser Zeit Laramidia nennen, entdeckt. Sicher belegt ist er aus der Hell Creek-, Lance-, Evanston– und Denver Formation aus den USA sowie aus der Scollard– und Frenchman Formation von Kanada. Einige Funde, die jedoch bislang nicht klar zugeordnet werden können, stammen auch aus der Ferris-, Hoppers-, Javelina– und Ojo Alamo Formation, sowie nahezu allen anderen für Dinosaurier-Fossilien bekannten Lagerstätten des westlichen Nordamerika.1Pearson, D. A. et al. (2002). Vertebrate biostratigraphy of the Hell Creek Formation in southwestern North Dakota and northwestern South Dakota. In Hartman, J. H., Johnson, K. R. & Nichols, D. J. (Hgg.), The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An integrated continental record of the end of the Cretaceous. Geological Society of America, 154. doi:10.1130/0-8137-2361-2.145 Fußspuren, die sehr wahrscheinlich zu Triceratops gehören, sind außerdem aus der Laramie Formation bekannt.2Tweet, J. (2025). Paleontology of Morrison–Golden Fossil Areas National Natural Landmark. Park Paleontology News, 17(2). Triceratops lebte somit sicher in den blauen, hellroten und auch violetten Regionen, also den Gestaden des Binnenmeeres, dem milden Nordwesten und dem westlichen Hochland. Möglicherweise kam er aber deutlich weiter südlich vor, sodass sein Verbreitungsgebiet bis zu den Küsten im Südwesten (gelb) reichte. Damit wäre er einer der am weitesten verbreiteten Dinosaurier Laramidias. |
Forschungsgeschichte
 George L. Cannon George L. Cannon |
Das erste benannte Fossil, das heute Triceratops zugeschrieben wird, war ein Paar imposanter Hörner am Teil eines Schädeldachs, die 1887 von George L. Cannon nahe Denver in Colorado, gefunden wurden. |
 Othniel C. Marsh Othniel C. Marsh |
Das Exemplar wurde an den berühmten Paläontologen Othniel C. Marsh geschickt, der annahm, die Gesteinsschicht stamme aus dem Pliozän und die Knochen gehörten zu einem besonders großen und ungewöhnlichen Bison, den er Bison alticornis nannte.3Marsh, O. C. (1887). Notice of new fossil mammals. American Journal of Science, 34(202), 323–331. doi:10.2475/ajs.s3-34.202.323 Bereits im folgenden Jahr erkannte Marsh jedoch anhand eines weiteren Exemplars, dass es gehörnte Dinosaurier gab, und veröffentlichte die Gattung Ceratops auf Grundlage fragmentarischer Überreste. Dabei stellte er auch die Familie Ceratopsidae auf.4Marsh, O. C. (1888). A new family of horned Dinosauria, from the Cretaceous. American Journal of Science, 36(216), 477–478. doi:10.2475/ajs.s3-36.216.477 |
 John B. Hatcher John B. Hatcher |
Ein drittes Exempar entdeckte 1888 ein Cowboy namens Edmund B. Wilson inmitten einer steilen Schluchtwand in der Lance Formation in Wyoming. Als er versuchte, den riesigen Schädel mit seinem Lasso an einem der Hörner herauszuziehen, brach das Horn ab und der Schädel stürzte in die Tiefe. Wilson brachte das Horn zu seinem Arbeitgeber, dem Rancher und Fossiliensammler Charles A. Guernsey, der es zufällig dem bekannten Knochenjäger John B. Hatcher zeigte. |
Hatcher konnte den gefallenen Schädel im Auftrag von Othniel C. Marsh bergen und schickte das Fossil nach Yale. Marsh benannte ihn dort zunächst als eine weitere Art von Ceratops, C. horridus. Nachdem bei der weiteren Präparation ein drittes Horn, nämlich das auf der Nase freigelegt wurde, änderte Marsh den Namen zu Triceratops („Dreihorngesicht“).5Marsh, O. C. (1889a). Notice of new American Dinosauria. American Journal of Science, 37(220), 331–336. doi:10.2475/ajs.s3-37.220.3316Marsh, O. C. (1889b). Notice of gigantic horned Dinosauria from the Cretaceous. American Journal of Science, 38(224), 173–175. doi:10.2475/ajs.s3-38.224.173

 Edward D. Cope Edward D. Cope |
Doch war Marsh wirklich der erste, der sich mit Triceratops wissenschaftlich befasste? Wahrscheinlich nicht! Denn bereits 1872 und 1874 hatte Marshs Erzrivale Edward D. Cope Fossilien beschrieben, die sehr wahrscheinlich zu diesem Hornsaurier gehörten. Unter den Namen Agathaumas sylvestris und Polyonax mortuarius hatte er sogar zwei Gattungen aufgestellt.7Cope, E. D. (1872). On the existence of Dinosauria in the Transition Beds of Wyoming. Proceedings of the American Philosophical Society, 12, 481–483.8Cope, E. D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. Bulletin of the U.S. Geological and Geographical Survey of the Territories, 9, 9–28. |
Agathaumas beruhte auf einem Becken, mehreren Wirbeln und einigen Rippen, die nahe dem Green River im Südosten Wyomings aus Schichten der maastrichtischen Lance Formation geborgen worden waren.9Lull, R. S. & Wright, N. E. (1942). Hadrosaurian dinosaurs of North America (Vol. 40). Geological Society of America. Aufgrund der starken Fragmentierung lassen sich diese Überreste jedoch nur sicher der Familie Ceratopsidae zuordnen.10Dodson, P. (1996). The horned dinosaurs. Princeton University Press.11Lull, R. S. (1933). A revision of the Ceratopsia or horned dinosaurs. Memoirs of the Peabody Museum of Natural History, 3(3), 1–175. doi:10.5962/bhl.title.5716 Polyonax mortuarius entdeckte Cope 1873 selbst im Nordosten Colorados, möglicherweise in der Denver Formation. Das Material bestand lediglich aus fragmentarischen Hornzapfen, drei Rückenwirbeln und Teilen von Gliedmaßenknochen.12Cope, E.D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. Bulletin of the U.S. Geological and Geographical Survey of the Territories. 9:9-28. Wie bei Agathaumas erlaubt auch hier der Erhaltungszustand keine genauere Zuordnung über die Ceratopsidae hinaus.13Dodson, P., Forster, C. A. & Sampson, S. D. (2004). Ceratopsidae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Eds.), The dinosauria. University of California Press, 494–513.
Schon im 19. Jahrhundert wurden über dreißig weitere Triceratops und bis heute mehrere hundert Exemplare, darunter auch etwa 120 gut erhaltene Schädel entdeckt. So ist dieser Dinosaurier inzwischen in den US-Bundesstaaten Colorado, Montana, North Dakota, South Dakota, Texas, Utah und Wyoming sowie aus den kanadischen Provinzen Saskatchewan und Alberta bekannt. Beinahe ein Drittel aller gefundenen Dinosaurierfossilien aus dem oberen Maastrichtium Nordamerikas können Triceratops inzwischen zugerechnet werden. Damit ist er der häufigste Dinosaurier seiner Zeit, der erfolgreichste und bekannteste Horndinosaurier (Ceratopsidae) überhaupt, obwohl die meisten Exemplare nur aus Teilskeletten oder isolierten Schädeln bestehen.

Taxonomische Debatten: Was ist eigentlich ein Triceratops, und was nicht?
Im Laufe der langen Zeit hat man diese auch vielen verschiedenen Arten von Triceratops oder auch ganz anderen Gattungen zugeordnet.14Hatcher, J. B., Marsh, O. C. & Lull, R. S. (1907). The Ceratopsia. Government Printing Office. So gehören nicht nur Copes Agathaumas und Polyonax sehr wahrscheinlich zu Triceratops, sondern auch die Gattungen Nedoceratops15Mateus, O. (2008). Two ornithischian dinosaurs renamed – Microceratops Bohlin 1953 and Diceratops Lull 1905. Journal of Paleontology, 82(2), 423. doi:10.1666/07-069.116Scannella, J.; Horner, J.R. (2010). „Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny“. Journal of Vertebrate Paleontology. 30 (4): 1157–1168. Bibcode:2010JVPal..30.1157S. doi:10.1080/02724634.2010.483632. S2CID 86767957.17Farke, A. A. (2011). Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE, 6(1), e16196. doi:10.1371/journal.pone.0016196, Tatankaceratops18Ott, C. J. & Larson, P. L. (2010). A new, small ceratopsian dinosaur from the Latest Cretaceous Hell Creek Formation, northwest South Dakota, United States – a preliminary description. In Ryan, M. J., Chinnery-Allgeier, B. J. & Eberth, D. A. (Eds.), New perspectives on horned dinosaurs – the Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press, 656.19Longrich, N. R. (2011). Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretaceous Research, 32(3), 264–276. doi:10.1016/j.cretres.2010.12.00720Holtz, T. R. Jr. (2011). Dinosaurs – the most complete, up-to-date encyclopedia for dinosaur lovers of all ages, Winter 2010 appendix.21Illies, M. M. et al. (2020). Triceratops with a kink – co-ossification of five distal caudal vertebrae from the Hell Creek Formation of North Dakota. Cretaceous Research, 108, 104355.22Scannella, J. & Horner, J. R. (2010). Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae) – synonymy through ontogeny. Journal of Vertebrate Paleontology, 30(4), 1157–1168. doi:10.1080/02724634.2010.483632 und Ojoceratops.23Sullivan, R. M. & Lucas, S. G. (2006). The Kirtlandian land-vertebrate „age“–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America. New Mexico Museum of Natural History and Science Bulletin, 35, 7–29.24Jasinski, S. E., Sullivan, R. M. & Dodson, P. (2020). New dromaeosaurid dinosaur (Theropoda, Dromaeosauridae) from New Mexico and biodiversity of dromaeosaurids at the end of the Cretaceous. Scientific Reports, 10(1), 5105. doi:10.1038/s41598-020-61480-725Sullivan, R. M. & Lucas, S. G. (2010). A new chasmosaurine (Ceratopsidae, Dinosauria) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. In Ryan, M. J., Chinnery-Allgeier, B. J. & Eberth, D. A. (Eds.), New perspectives on horned dinosaurs – The Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press.26Longrich, N. R. (2011). Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretaceous Research, 32(3), 264–276. doi:10.1016/j.cretres.2010.12.00727Holtz, T. R. Jr. (2011). Dinosaurs – the most complete, up-to-date encyclopedia for dinosaur lovers of all ages, Winter 2010 appendix.28Jasinski, S. E., Sullivan, R. M. & Lucas, S. G. (2011). Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member) San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science Bulletin, 53, 216–271.29Maiorino, L. et al. (2015). Variation in the shape and mechanical performance of the lower jaws in ceratopsid dinosaurs (Ornithischia, Ceratopsia). Journal of Anatomy, 227(5), 631–646. doi:10.1111/joa.1237430Brown, C. M. & Henderson, D. M. (2015). A new horned dinosaur reveals convergent evolution in cranial ornamentation in Ceratopsidae. Current Biology, 25(12), 1641–1648. doi:10.1016/j.cub.2015.04.041

 John Scanella John Scanella
|
Auch Torosaurus latus könnte nach Auffassung einiger Wissenschaftler ein Synonym von Triceratops sein. Unter anderem interpretierten John Scanella und Jack Horner dieses Tier mit seinem verlängerten Nackenschild mit den markanten beiden Fenstern als ein voll ausgewachsenes Individuum, wogegen die Triceratops-Funde mit kürzerem Nackenschild von subadulten Tieren stammen sollten. Im Laufe des Wachstums würde bei Triceratops das Knochengewebe der Nackenschilde immer dünner werden und sich schließlich ganz zurückbilden, wodurch bei sehr alten Individuen schließlich die beiden Fenster entstünden. 31Scannella, J. (2009). And then there was one – synonymy consequences of Triceratops cranial ontogeny. Journal of Vertebrate Paleontology, 29, 177A.32Scannella, J. & Horner, J. R. (2010). Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae) – synonymy through ontogeny. Journal of Vertebrate Paleontology, 30(4), 1157–1168. doi:10.1080/02724634.2010.483632 |


Diese Ansicht wird jedoch von vielen Kollegen angezweifelt. Die dünnen Knochenbereiche am Nackenschild können auch als Muskelansatzstellen erklärt werden. Zudem gebe es keinen konsistenten Zusammenhang zwischen Öffnungen und einer granulierten Knochenstruktur.33Farke, A. A. (2011). Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE, 6(1), e16196. doi:10.1371/journal.pone.0016196
 Nick Longrich Nick Longrich |
Auch Nick Longrich befasste sich 2012 mit der sogenannten „Toromorph-Hypothese“ und formulierte drei überprüfbare Vorhersagen: Erstens müssten Triceratops und Torosaurus in denselben Regionen vorkommen, was jedoch nicht vollständig zutrifft. Zweitens dürften nur Torosaurus-Exemplare vollständig ausgewachsen sein, während es alte Triceratops überhaupt nicht geben dürfte. Allerdings existieren sowohl Hinweise für jugendliche Torosaurus– als auch sehr alte Triceratops-Schädel. Drittens müssten auch klare Übergangsformen existieren. Doch die angeblichen Vorstufen der Schädelfenster unterscheiden sich anatomisch deutlich, weshalb Longrich die Toromorph-Hypothese verwarf.34Longrich, N. R. & Field, D. J. (2012). Torosaurus is not Triceratops – ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy. PLoS ONE, 7(2), e32623. doi:10.1371/journal.pone.0032623 |
Im Jahr 2022 wurden zudem zwei subadulte Exemplare aus der Frenchman Formation und der Scollard Formation Kanadas beschrieben und konnten eindeutig Torosaurus zugeordnet werden. Dies stützt die Auffassung, dass Torosaurus ein gültiges, eigenständiges Taxon ist und bis in das späte Maastrichtium existierte.35Mallon, J. C. et al. (2022). The record of Torosaurus (Ornithischia: Ceratopsidae) in Canada and its taxonomic implications. Zoological Journal of the Linnean Society, 195(1), 157–171. doi:10.1093/zoolinnean/zlab120 Sollte sich eines Tages jedoch herausstellen, dass Ojoceratops oder Torosaurus doch mit Triceratops synonym sind, würde dies das Verbreitungsgebiet von Triceratops auch nach New Mexico ausweiten.
Während meiner Recherchearbeit zu Die weißen Steine war der Disput um Torosaurus als möglicher Triceratops noch in vollem Gange. Da mich die Ideen von John Scanella und Jack Horner zwar faszinierten, aber nicht völlig überzeugten, wagte ich mich stattdessen an eine eigene These: Torosaurus ist in meinen Romanen das (durch Geschlechtsdimorphismus etwas kleinere!) Männchen von Triceratops. Da Torosaurus-Fossilien außerdem seltener gefunden wurden als die des größeren Triceratops, lebt ein ausgewachsenes Triceratops-Männchen in meiner Welt auch eher einzelgängerisch, während die Weibchen Herden bilden. Möglicherweise wurden sie deshalb häufiger von Fressfeinden erbeutet oder hielten sich längere Zeit in zur Fossilisation weniger gut geeigneten Gegenden auf, sodass sie im Fossilbericht seltener auftauchen. Das ist aber wie gesagt nur eine Spekulation, genau wie das im Buch beschriebene Wanderverhalten, mit der ich die geringere Fossiliendichte von „meinem“ Triceratops (Torosaurus) erklären möchte.
Abgesehen von den geschilderten Problematiken werden von Triceratops heute bloß noch zwei Arten als gültig anerkannt: Triceratops horridus und Triceratops prorsus. Erstere scheint geologisch etwas älter zu sein und kommt in den unteren Schichten der Hell Creek Formation vor. Die zweite ist dagegen nur aus Schichten bekannt, die unmittelbar vor der KP-Grenze liegen. Es handelt sich also wahrscheinlich um aufeinanderfolgende Chronospezies: zu unterschiedlichen Zeiten lebende Vertreter von ein und derselben Stammlinie, so wie etwa bei uns Menschen Homo erectus und Homo sapiens.36Scannella, J. B. & Fowler, D. W. (2009). Anagenesis in Triceratops – evidence from a newly resolved stratigraphic framework for the Hell Creek Formation. 9th North American Paleontological Convention Abstracts. Cincinnati Museum Center Scientific Contributions, 3, 148–149.37Mallon, J. C. et al. (2025). The Canadian fossil record supports anagenesis in Triceratops (Ornithischia, Ceratopsia). Canadian Journal of Earth Sciences, 62(7), 1222–1236. doi:10.1139/cjes-2024-0170

Weil Triceratops horridus viel bekannter ist als Triceratops prorsus, habe ich in meiner ersten Buchfassung zuerst nur den T. horridus auftreten lassen. Die These mit den verschiedenen Chronospezies kannte ich damals noch nicht. Da Die Weißen Steine aber in der Zeit des obersten Maastrichtiums spielt, müsste dort eigentlich nur T. prorsus vorkommen. Dieser Fehler wurde in der Neuauflage korrigiert.
Systematik und Verwandtschaft
Unabhängig davon, ob Taxa wie Torosaurus, Nedoceratops oder Tatankaceratops nun eigene Gattungen repräsentieren, ist auch die genaue systematische Stellung sowie die Abstammung von Triceratops innerhalb der Ceratopsiden umstritten.
 Lawrence M. Lambe Lawrence M. Lambe |
Die spätkreidezeitlichen Horndinosaurier wurden schon 1915 von Lawrence M. Lambe in zwei große Gruppen eingeteilt: einerseits die Centrosaurinae mit kurzem, aber oft durch auffällige Stacheln und Anhängsel verzierten Nackenschildern, einem meist massiven Nasenhorn oder breitem Nasenkamm sowie kurzen bzw. ganz fehlenden Stirnhörnern. Ihnen stehen auf der anderen Seite die Chasmosaurinae gegenüber, die wiederum nur sehr kleine Nasenhörnern, dafür aber mächtige Stirnhörner und einen sehr großen Nackenschild mit auffälligen Fenstern besitzen.38Lambe, L. M. (1915). On Eoceratops canadensis, gen. nov., with remarks on other genera of Cretaceous horned dinosaurs. Museum Bulletin No. 12, Geological Series No. 24. Geological Survey, Department of Mines, Ottawa.39Madzia, D. et al. (2021). The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ, 9, e12362. doi:10.7717/peerj.12362 |

Triceratops vereint Merkmale beider Gruppen, da er sowohl sehr lange Brauenhörner, aber einen verhältnismäßig kurzen, massiven Nackenschild besitzt. Somit wurde er zunächst als Vertreter der Centrosaurinen interpretiert.40Hatcher, J. B., Marsh, O. C. & Lull, R. S. (1907). The Ceratopsia. Government Printing Office, Washington, D.C.41Lull, R. S. (1933). A revision of the Ceratopsia or horned dinosaurs. Memoirs of the Peabody Museum of Natural History, 3(3), 1–175. doi:10.5962/bhl.title.5716
 Charles M. Sternberg Charles M. Sternberg |
1949 stellte Charles M. Sternberg diese Einordnung erstmals infrage. Aufgrund verschiedener Schädel- und Hornmerkmale argumentierte er, dass Triceratops näher mit Arrhinoceratops und Chasmosaurus verwandt sei und somit zu den Chasmosaurinen gehöre.42Sternberg, C. M. (1949). The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member. Phylogeny of the Ceratopsidae. National Museum of Canada Bulletin, 113, 33–46. |
Seine Ansicht wurde zunächst weitgehend ignoriert, da die meisten Fachkollegen auch noch bis in die 80er Jahre des 20. Jahrhunderts weiterhin eine Zugehörigkeit zu den Centrosaurinen annahmen.43Ostrom, J. H. (1966). Functional morphology and evolution of the ceratopsian dinosaurs. Evolution, 20(3), 290–308. doi:10.2307/2406631 44Norman, D. (1985). The Illustrated Encyclopaedia of Dinosaurs. Salamander Books, London. Spätere Funde und Analysen, insbesondere von Thomas Lehmann und Peter Dodson, bestätigten jedoch Sternbergs Einschätzung, und seither gilt Triceratops als Vertreter der Chasmosaurinae.
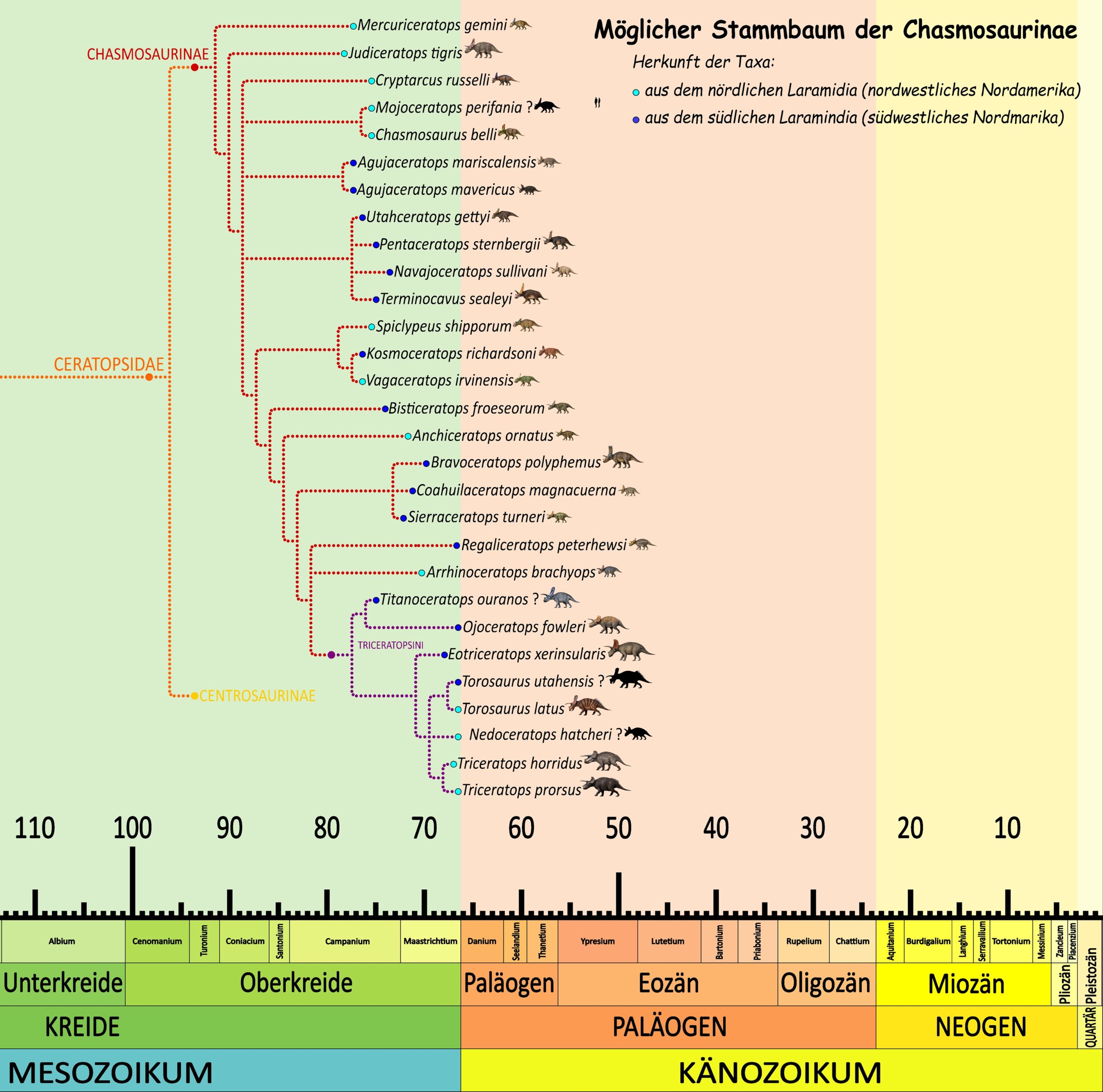
Die entwicklungsgeschichtliche Herkunft von Triceratops ist jedoch nach wie vor unklar. 1922 hielt man den damals neu entdeckten Protoceratops für den ältesten Vorfahren von Triceratops.45Granger, W. W. & Gregory, W. K. (1923). Protoceratops andrewsi, a pre-ceratopsian dinosaur from Mongolia. American Museum of Natural History Novitates, 72, 1–9. Erst viele Jahrzehnte später wurden weitere wichtige und auch weit ältere Übergangsformen entdeckt. Besonders bedeutend waren Zuniceratops, der früheste bekannte Ceratopsier mit Brauenhörnern,46Wolfe, D. G. & Kirkland, J. I. (1998). Zuniceratops christopheri n. gen. & n. sp., a ceratopsian dinosaur from the Moreno Hill Formation (Cretaceous, Turonian) of west-central New Mexico. New Mexico Museum of Natural History and Science, Bulletin, 14, 303–317. sowie Yinlong, der erste bekannte jurassische Ceratopsier überhaupt.47Xu, X., Forster, C. A., Clark, J. M. & Mo, J. (2006). A basal ceratopsian with transitional features from the Late Jurassic of northwestern China. Proceedings of the Royal Society B: Biological Sciences, 273(1598), 2135–2140. doi:10.1098/rspb.2006.3566 Diese Entdeckungen lieferten wichtige Hinweise auf die Herkunft der Ceratopsier insgesamt. Heute geht man davon aus, dass die Gruppe ihren Ursprung in Asien während des Jura hatte und sich die „echten“ gehörnten Ceratopsier erst zu Beginn der Oberkreide in Nordamerika entwickelten. Triceratops gehörte schließlich zu den letzten Ceratopsiern überhaupt, war einer ihrer größten und massigsten Vertreter und sollte während des großen Massenaussterbens vor 66 Millionen Jahren zusammen mit allen anderen Dinosauriern aussterben.

Auch in der phylogenetischen Systematik der Dinosaurier spielt Triceratops eine wichtige Rolle: Die Dinosauria werden heute als alle Nachkommen des letzten gemeinsamen Vorfahren von Triceratops und den heutigen Vögeln definiert.48Gauthier, J. A. (1986). Saurischian monophyly and the origin of birds. The Origin of Birds and the Evolution of Flight, K. Padian (Hg.). Memoirs of the California Academy of Sciences, 8, 1–55. Ebenso werden die Ornithischia als jene Dinosaurier definiert, die näher mit Triceratops verwandt sind als mit modernen Vögeln.49Sereno, P. C. (1998). A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 210(1), 41–83. doi:10.1127/njgpa/210/1998/41
Körperbau
Triceratops war ein sehr großes Tier, vergleichbar von seiner Masse mit einem heutigen afrikanischen Elefanten. Er erreichte eine Länge von etwa 8 bis 9 Metern sowie ein Gewicht von ungefähr 6 bis 10 Tonnen.50Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press, 265–267. Wie alle Vertreter der Chasmosaurinen besaß Triceratops einen im Verhältnis zum Körper enorm großen Schädel und einen der größten aller bekannten Landtiere überhaupt. Der größte bekannte Schädel, Exemplar MWC 7584 (früher BYU 12183), wird vollständig rekonstruiert auf etwa 2,5 Meter Länge geschätzt und machte damit fast ein Drittel der gesamten Körperlänge des Tieres aus.51Lambert, D. (1993). The Ultimate Dinosaur Book. Dorling Kindersley, New York. pp. 152–167. ISBN 978-1-56458-304-8.
Die Unterkiefer waren langgestreckt und trafen sich vorne in einem gemeinsamen Epidentale, dem Kern des zahnlosen unteren Schnabels. Im hinteren Teil des Kiefers verliefen Zahnbatterien mit 36 bis 40 Zahnpositionen, wobei pro Position drei bis fünf Zähne vertikal übereinander standen. Insgesamt verfügte Triceratops somit über etwa 432 bis 800 Zähne, von denen jedoch immer nur ein Teil gleichzeitig aktiv genutzt wurde, da die Zähne lebenslang ersetzt wurden. Triceratops war ein Pflanzenfresser. Aufgrund seines tief gehaltenen Kopfes bestand seine Nahrung wahrscheinlich hauptsächlich aus niedrig wachsender Vegetation. Dennoch könnte das Tier mit seinen Hörnern, dem kräftigen Schnabel und seiner enormen Körpermasse auch höhere Pflanzen umgestoßen haben.52Tait, J. & Brown, B. (1928). How the Ceratopsia carried and used their head. Transactions of the Royal Society of Canada, 22, 13–23. Die Kiefer endeten in einem tiefen, schmalen Schnabel, der vermutlich besser zum Greifen und Abreißen von Pflanzen geeignet war als zum eigentlichen Beißen.53Ostrom, J. H. (1966). Functional morphology and evolution of the ceratopsian dinosaurs. Evolution, 20(3), 290–308. doi:10.2307/240663154Erickson, G. M. et al. (2015). Wear biomechanics in the slicing dentition of the giant horned dinosaur Triceratops. Science Advances, 1(5), e1500055. doi:10.1126/sciadv.1500055 Die Zahnbatterien arbeiteten anschließend wie eine große Schere: Die Zähne bewegten sich nahezu vertikal gegeneinander und konnten das abgerissene Pflanzenmaterial effektiv zerkleinern. Beim Fressen nutzten sich die Zähne außerdem so ab, dass sich längliche Vertiefungen bildeten, die die Reibung beim Kauen verringerten. Die enorme Körpergröße und die Vielzahl der Zähne deuten darauf hin, dass Triceratops große Mengen faserreicher Pflanzen fraß. Zu den möglichen Nahrungspflanzen gehörten Vertreter der Pappeln (Populus), Kiefern, Platanen, Haselgewächse sowie Sumpfzypressen (Taxodium). Einige Forscher vermuten außerdem, dass Triceratops ebenso wie sein naher Verwandter Torosaurus Zykadeen und Palmgewächse fraß, während andere eher Farne als Hauptnahrung annehmen, die damals in offenen Ebenen wuchsen.55Maiorino, L., Farke, A. A., Kotsakis, T., Teresi, L. & Piras, P. (2015). Variation in the shape and mechanical performance of the lower jaws in ceratopsid dinosaurs (Ornithischia, Ceratopsia). Journal of Anatomy, 227(5), 631–646. doi:10.1111/joa.1237456Coe, M. J., Dilcher, D. L., Farlow, J. O., Jarzen, D. M. & Russell, D. A. (1987). Dinosaurs and land plants. In Friis, E. M., Chaloner, W. G. & Crane, P. R. (Hrsg.), The Origins of Angiosperms and their Biological Consequences. Cambridge University Press, 225–258.57de Rooij, J. et al. (2022). Stable isotope record of Triceratops from a mass accumulation (Lance Formation, Wyoming, USA) provides insights into Triceratops behaviour and ecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 607, 111274. doi:10.1016/j.palaeo.2022.111274 Isotopenuntersuchungen an den Zähnen von Ceratopsiern und Hadrosauriern zeigen zudem, dass Triceratops und Edmontosaurus wahrscheinlich unterschiedliche ökologische Nahrungsquellen bevorzugten. Dadurch konnten beide großen Pflanzenfresser im selben Lebensraum existieren, ohne miteinander zu konkurrieren.58Fricke, H. C. & Pearson, D. A. (2008). Stable isotope evidence for changes in dietary niche partitioning among hadrosaurian and ceratopsian dinosaurs of the Hell Creek Formation, North Dakota. Paleobiology, 34(4), 534–552. doi:10.1666/08020.159Sander, P. M., Gee, C. T., Hummel, J. & Clauss, M. (2010). Mesozoic plants and dinosaur herbivory. In Gee, C. T. (Hg.), Life of the Past, 331–359. doi:10.5167/uzh-3528360Barrett, P. M. & Rayfield, E. J. (2006). Ecological and evolutionary implications of dinosaur feeding behaviour. Trends in Ecology & Evolution, 21(4), 217–224. doi:10.1016/j.tree.2006.01.002

Der Gelenkknochen am hinteren Ende des Unterkiefers war außergewöhnlich breit und entsprach der allgemeinen Breite des Kiefergelenks. Über den Nasenöffnungen saß ein einzelnes Horn auf der Schnauze, zwei weitaus größere Brauenhörner saßen auf der Stirn. Bei einigen Exemplaren waren sie über einen Meter lang. Am hinteren Teil des Schädels besaß Triceratops einen im Vergleich zu anderen Chasmosaurinen relativ kurzen knöchernen Nackenschild. Bei jüngeren Tieren war dieser mit sogenannten Epoccipitalia versehen, also dreieckigen Knochenfortsätzen am Rand des Schildes. Die meisten anderen Ceratopsiden besaßen große Öffnungen im Nackenschild, sogenannte Parietalfenster. Bei Triceratops war der Schild dagegen auffallend massiv und geschlossen. Unterhalb des Nackenschildes befand sich am Hinterkopf ein riesiger Hinterhauptskondylus mit bis zu 106 Millimetern Durchmesser, der eine hohe Beweglichkeit des Kopfes erlaubte. 61Dodson, P., Forster, C. A. & Sampson, S. D. (2004). Ceratopsidae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hrsg.), The Dinosauria. University of California Press, Berkeley, 494–513.62Scannella, J. B. et al. (2014). Evolutionary trends in Triceratops from the Hell Creek Formation, Montana. Proceedings of the National Academy of Sciences, 111(28), 10245–10250. doi:10.1073/pnas.1313334111


Die Wirbelsäule bestand aus zehn Halswirbeln, zwölf Rückenwirbeln, zehn Kreuzbeinwirbeln und etwa fünfundvierzig Schwanzwirbeln. Das Becken war groß und besaß ein langes Darmbein (Ilium), das Sitzbein (Ischium) war nach unten gebogen. Insgesamt war das Skelett von Triceratops kaum anders gebaut als das seiner Verwandten, aber ausgesprochen robust. Er besaß einen massigen Körper mit kräftigen Gliedmaßen, kurzen Händen mit jeweils drei hufartigen Fingern sowie kurzen Füßen mit vier Zehen, ebenfalls mit hufartigen Enden. Die Vorderbeine von Triceratops bewahrten im Vergleich zu anderen vierbeinigen Dinosauriern eine relativ ursprüngliche Struktur. Bei vielen Thyreophoren und Sauropoden waren sie so gedreht, dass die Handflächen nach hinten zeigten. Bei Triceratops und anderen Ceratopsiern sowie bei den verwandten vierbeinigen Ornithopoden zeigten sie beim Laufen aber seitlich nach außen. Das Körpergewicht wurde bei Triceratops nur von den ersten drei Fingern getragen. Der vierte und fünfte Finger waren stark reduziert und besaßen weder Krallen noch Hufe. Obwohl Triceratops eindeutig vierbeinig lief, wurde seine Körperhaltung lange diskutiert. Früher nahm man an, die Vorderbeine hätten seitlich abgespreizt gestanden, um das enorme Gewicht des Kopfes besser tragen zu können. Spurenfossilien sowie moderne Rekonstruktionen physischer und digitaler Skelette zeigen jedoch, dass Triceratops und andere Ceratopsier wahrscheinlich eine aufrechte Haltung einnahmen. Die Ellenbogen waren nach hinten gebogen und nur leicht nach außen gerichtet.63Christiansen, P. & Paul, G. S. (2001). Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs. Gaia, 16, 13–29.64Thompson, S. & Holmes, R. (2007). Forelimb stance and step cycle in Chasmosaurus irvinensis (Dinosauria: Neoceratopsia). Palaeontologia Electronica, 10(1), 17.65Fujiwara, S.-I. (2009). A reevaluation of the manus structure in Triceratops (Ceratopsia: Ceratopsidae). Journal of Vertebrate Paleontology, 29(4), 1136–1147. doi:10.1671/039.029.040666Rega, E., Holmes, R. & Tirabasso, A. (2010). Habitual locomotor behavior inferred from manual pathology in two Late Cretaceous chasmosaurine ceratopsid dinosaurs, Chasmosaurus irvinensis (CMN 41357) and Chasmosaurus belli (ROM 843). In Ryan, M. J., Chinnery-Allgeier, B. J. & Eberth, D. A. (Hgg.), New Perspectives on Horned Dinosaurs. The Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press, Bloomington and Indianapolis, 340–354.
Sozialverhalten und Häufigkeit
Obwohl Triceratops fast in allen populären Darstellungen als Herdentier dargestellt wird, gab es bis vor kurzem kaum Hinweise darauf, dass diese Dinosaurier tatsächlich gesellig lebten. Da von mehreren anderen Ceratopsiern Knochenlagerstätten mit Überresten von Dutzenden bis sogar Tausenden Individuen bekannt sind, wurde die dort recht naheliegende gesellige Lebensweise einfach auf Triceratops übertragen. Erst seit kurzer Zeit weiß man aber, dass dies wahrscheinlich auch zutraf. 2009 wurde im Südosten Montanas ein Bonebed beschrieben, das überwiegend aus Triceratops-Knochen besteht, von insgesamt drei Jungtieren.67Mathews, J. C., Brusatte, S. L., Williams, S. A. & Henderson, M. D. (2009). The first Triceratops bonebed and its implications for gregarious behavior. Journal of Vertebrate Paleontology, 29(1), 286–290. doi:10.1080/02724634.2009.10010382 2012 wurden nahe Newcastle in Wyoming zwei relativ vollständig erhaltene ausgewachsene Triceratops-Individuen zusammen mit einem Jungtier entdeckt. Die Knochen zeigen zudem deutliche Spuren von Angriffen bzw. Aasfraß durch Tyrannosaurus, besonders am größten Exemplar.68Smith, M. (2013). Triceratops trio unearthed in Wyoming. CNN. Eine Studie aus dem Jahr 2020 beschrieb miteinander verwachsene Schwanzwirbel von Triceratops. Die Forscher vermuteten, dass diese Verletzung entstanden sein könnte, nachdem ein Tier versehentlich auf den Schwanz eines anderen Herdemitglieds getreten war.69Illies, M. M. C. & Fowler, D. W. (2020). Triceratops with a kink. Co-ossification of five distal caudal vertebrae from the Hell Creek Formation of North Dakota. Cretaceous Research, 108, 104355. doi:10.1016/j.cretres.2019.104355 Den bisher stärksten Beleg für eine Lebensweise zumindest in kleinen Sozialverbänden lieferte eine Ausgrabung des Naturalis Biodiversity Center im niederländischen Leiden, die 2013 insgesamt fünf Individuen in Wyoming entdeckte. Nach über 10 Jahren Präparationsarbeit und intensiver Forschung wurden sie in einer Ausstellung präsentiert. Aktuelle Forschungsarbeiten dazu mit den Ergebnissen zur sozialen Lebensweise stehen offenbar kurz vor der Veröffentlichung.
 |
Triceratops war wahrscheinlich zu seiner Zeit einer der häufigsten Dinosaurier seines Lebensraums. So gehören Zähne, Hornfragmente, Teile des Nackenschildes und andere Schädelreste zu den häufigsten Fossilien von Dinosauriern, die man in den einschlägigen Fundgebieten finden kann. Robert T. Bakker schätzte 1986, dass Triceratops rund fünf Sechstel der großen Dinosaurierfauna am Ende der Kreidezeit ausmachte. |
Auffällig ist außerdem, dass Schädelreste wesentlich häufiger gefunden werden als Knochen des übrigen Körpers. Dies deutet darauf hin, dass die massiven Schädel besonders gute Fossilisationsbedingungen besaßen. Eine andere Erklärung: egal, auf welche Weise ein Triceratops starb, ob nach einem Kampf, an Krankheit oder sonstigen Umständen, zog sein Kadaver wohl relativ schnell die Aufmerksamkeit eines Tyrannosaurus auf sich. Da dieser Fleischfresser auch Knochen zerbeißen und das Knochenmark verwenden konnte, aber den massiven Schädel wohl nicht, könnten vor allem die Schädel nach seinem Mahl als „Abfall“ übrig geblieben sein. Insbesondere um an die kräftigen und nahrhaften Hals- und Rückenmuskeln zu kommen, könnte Tyrannosaurus die toten Triceratops regelrecht geköpft haben. Tatsächlich weisen mehrere Schädel von Triceratops typische postmortale Verletzungen auf, die genau auf dieses Verhalten hinweisen.70Fowler, Denver & Scannella, John & Goodwin, Mark. (2012). How to eat a Triceratops: large sample of toothmarks provides new insight into the feeding behavior of Tyrannosaurus. J. Vert. Paleontol.. 32. 96-96. In Die Weißen Steine habe ich so eine Köfpung entsprechend authentisch beschrieben.
Sinnesleistungen und Stoffwechsel
Untersuchungen des Gehirnschädels deuten darauf hin, dass Triceratops im Vergleich zu vielen anderen Dinosauriern einen eher schwach entwickelten Geruchssinn hatte. Die Ohren waren dagegen vermutlich auf tieffrequente Geräusche spezialisiert,71Sakagami, R. & Kawabe, S. (2020). Endocranial anatomy of the ceratopsid dinosaur Triceratops and interpretations of sensory and motor function. PeerJ, 8, e9888. doi:10.7717/peerj.9888 ganz im Gegensatz zu früheren Ceratopsiern wie Yinlong, Liaoceratops und Psittacosaurus, die offenbar deutlich besser riechen und hören konnten als Triceratops.72Hu, J., Xu, X., Zhao, Q., He, Y., Forster, C. A. & Han, F. (2024). Endocranial morphology of three early-diverging ceratopsians and implications for the behavior and the evolution of the endocast in ceratopsians. Paleobiology, 50(3), 490–502. doi:10.1017/pab.2024.25 Eine Analyse ergab außerdem, dass das Tier den Kopf wahrscheinlich in einem Winkel von etwa 45 Grad zum Boden hielt. Dadurch wurden Hörner und Nackenschild besonders gut präsentiert, während gleichzeitig effizientes Grasen möglich blieb. Eine Studie aus dem Jahr 2026 untersuchte den großen Nasenbereich von Triceratops genauer. Dabei kamen die Forscher zu dem Schluss, dass das Tier wahrscheinlich Nasenmuscheln besaß, ähnlich wie viele heutige Tiere. Beim Einatmen verdunstete dort Feuchtigkeit, wodurch Wärme abgegeben und das Blut gekühlt wurde, das Gehirn und Augen versorgte. Dies könnte wichtig gewesen sein, da der enorme und schwere Schädel sonst leicht überhitzt wäre. Die Studie zeigte außerdem, dass bei Ceratopsiern ein bestimmter Gesichtsnerv den Schnabel versorgte, während der sonst bei Reptilien übliche Verlauf durch Schnabel und Nasenhöhle blockiert war.73Tada, S. et al. (2026). Nasal soft-tissue anatomy of Triceratops and other horned dinosaurs. The Anatomical Record. doi:10.1002/ar.70150

Eine Untersuchung von 2022 kam zu dem Ergebnis, dass Triceratops wahrscheinlich einen ectothermen („kaltblütigen“) oder gigantothermen Stoffwechsel besaß, ähnlich wie große heutige Reptilien. Gigantothermie bedeutet, dass ein sehr großes Tier seine Körpertemperatur allein durch seine enorme Körpermasse relativ konstant halten kann. Andere Studien schließen jedoch nicht aus, dass Triceratops teilweise endotherme Eigenschaften besaß. Dieselben Untersuchungen zeigten außerdem, dass Triceratops in Überschwemmungsebenen und bewaldeten Binnenlandschaften lebte.74de Rooij, J. et al. (2022). Stable isotope record of Triceratops from a mass accumulation (Lance Formation, Wyoming, USA) provides insights into Triceratops behaviour and ecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 607, 111274. doi:10.1016/j.palaeo.2022.111274
Hautbeschaffenheit
Ein besonders bedeutender Fund ist ein Triceratops-Exemplar mit dem Spitznamen „Lane“ (HMNS PV.1506), das 2002 auf der Zerbst Ranch in Wyoming entdeckt wurde. Bei diesem Fossil sind große Bereiche der Haut über dem Rumpf erhalten geblieben. Sowohl das nahezu vollständige Skelett als auch die Hautfossilien werden heute im Houston Museum of Natural Science in Texas ausgestellt. Die Haut blieb nicht als bloßer Abdruck erhalten, sondern als dünne Tonschicht, die sich aus einem bakteriellen Film auf dem austrocknenden Körper bildete. Dadurch entstand ein außergewöhnlich detailreicher Positivabdruck der Hautoberfläche. Entgegen älteren Annahmen handelt es sich also nicht um echte konservierte Haut, sondern um eine extrem präzise mineralische Nachbildung ihrer Oberfläche.75Sereno, P. C. et al. (2026). Duck-billed dinosaur fleshy midline and hooves reveal terrestrial clay-template „mummification“. Science, 391(6780), eadw3536. doi:10.1126/science.adw3536

Die Schuppen besitzen kleine kegelförmige Erhebungen in ihrer Mitte. Dies führte zu der Hypothese, dass Triceratops möglicherweise borstenartige oder stachelige Hautstrukturen getragen haben könnte. Neuere Untersuchungen stellen diese Interpretation jedoch infrage. Zusätzlich ist ein erhaltenes Hautstück vom Nackenschild eines anderen Exemplars bekannt. Dieses zeigt kleine polygonale Grundschuppen, die eng aneinanderlagen.76Bell, P. R. et al. (2022). The exquisitely preserved integument of Psittacosaurus and the scaly skin of ceratopsian dinosaurs. Communications Biology, 5(1), 809. doi:10.1038/s42003-022-03749-3 Bei meiner Darstellung von Triceratops mit seinem doch etwas „struppigen“ Aussehen in Die weißen Steine habe ich mich an diesen neuen Erkenntnissen orientiert. Ob er aber wirklich so farbenprächtig war und auch ein Keratinüberzug über dem Nackenschild lag, ist rein spekulativ.
Funktion der Hörner und des Nackenschilds
In der Populärkultur ist Triceratops als kämpferischer Haudegen zur Legende geworden. Insbesondere die Szene, wie er seinen Widersacher Tyrannosaurus mit den Hörnern am Bauch aufspießt, ist eine sehr beliebte Darstellung.77Sternberg, C. H. (1917). Hunting dinosaurs in the Badlands of the Red Deer River, Alberta, Canada. C. H. Sternberg—The World Company Press. doi:10.5962/bhl.title.15981178Bakker, R. T. (1986). The dinosaur heresies – New theories unlocking the mystery of the dinosaurs and their extinction. William Morrow, 438 Dass die Hörner wirklich bei Kämpfen gegen Fressfeinde zum Einsatz kamen, ist durch eine verheilte Hornverletzung mit markanten T. rex-Bissmarken bei Triceratops zwar belegt,79Happ, J. (2008). An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. In Larson, P. & Carpenter, K. (Eds.), Tyrannosaurus rex, the Tyrant King (Life of the Past). Indiana University Press, 355–368. und auch, dass Tyrannosaurus nachweislich aktiv auf Triceratops Jagd machte.80Erickson, G. M. & Olson, K. H. (1996). Bite marks attributable to Tyrannosaurus rex – preliminary description and implications. Journal of Vertebrate Paleontology, 16(1), 175–178. doi:10.1080/02724634.1996.10011297 Doch zielte Triceratops dabei wohl eher auf die Beine oder die Flanken seines Feindes statt auf den Unterleib. Hätte Triceratops nämlich versucht, wie ein Kampfstier einen Tyrannosaurus auf die Hörner zu nehmen, so wäre der schwer verletzte Gegner auf seinen Kopf gefallen. Dabei hätte der sterbende T. rex wiederum den Triceratops durch sein tonnenschweres Gewicht schwer verletzen können.
.jpg)
Es ist inzwischen auch belegt, dass Triceratops seine Hörner und den mächtigen Nackenschild nicht nur zur Verteidigung gegen Raubtiere, sondern auch bei Kämpfen mit Artgenossen einsetzte. Eine Untersuchung von 2009 zeigte, dass die gefundenen Exemplare von Triceratops im Vergleich zu Centrosaurus deutlich häufiger Schädelverletzungen aufweisen. Etwa 14 % der untersuchten Exemplare zeigten entsprechende Schäden.81Farke, A. A. et al. (2009). Evidence of combat in Triceratops. PLoS ONE, 4(1), e4252. doi:10.1371/journal.pone.0004252 Histologische Befunde belegen zudem eine gute Heilungsfähigkeit des Nackenschildes.82Reid, R. E. H. (1997). Histology of bones and teeth. In Currie, P. J. & Padian, K. (Eds.), Encyclopedia of dinosaurs. Academic Press, 329–339.83Horner, J. R. & Lamm, E. (2011). Ontogeny of the parietal frill of Triceratops – a preliminary histological analysis. Comptes Rendus Palevol, 10(5–6), 439–452. doi:10.1016/j.crpv.2011.04.00684Farlow, J. O. & Dodson, P. (1975). The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. Evolution, 29(2), 353–361. doi:10.2307/2407222 Einzelne vernarbte Verletzungen, bei denen der Schild sogar vollständig durchstoßen wurde, wie etwa beim sehr großen Exemplar „Big John“, gelten als weitere starke Hinweise auf innerartliche Kämpfe.85D’Anastasio, R. et al. (2022). Histological and chemical diagnosis of a combat lesion in Triceratops. Scientific Reports, 12(1), 3941. doi:10.1038/s41598-022-08033-2 Neben einer möglichen Schutzfunktion des Schildes wurde auch Thermoregulation (ähnlich wie bei Elefantenohren) diskutiert,86Wheeler, P. E. (1978). Elaborate CNS cooling structures in large dinosaurs. Nature, 275(5679), 441–443. doi:10.1038/275441a0 doch die große Formenvielfalt der Ceratopsier spricht eher für eine wichtige Rolle des Nackenschildes in visueller Kommunikation, sozialer Interaktion und möglicherweise sexueller Selektion oder auch der Arterkennung.87Davitashvili, L. Sh. (1961). Teoriya Polovogo Otbora (Theory of Sexual Selection). Izdatel’stvo Akademii nauk SSSR, 538.88Farlow, J. O. & Dodson, P. (1975). The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. Evolution, 29(2), 353–361. doi:10.2307/240722289Lehman, T. M. (1990). The ceratopsian subfamily Chasmosaurinae – sexual dimorphism and systematics. In Carpenter, K. & Currie, P. J. (Eds.), Dinosaur Systematics – Perspectives and Approaches. Cambridge University Press, 211–229.90Forster, C. A. (1990). The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny. Ph.D. Dissertation thesis, University of Pennsylvania, 227.91Dodson, P., Forster, C. A. & Sampson, S. D. (2004). Ceratopsidae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Eds.), The dinosauria. University of California Press, 494–513.92Goodwin, M. B. et al. (2006). The smallest known Triceratops skull – new observations on ceratopsid cranial anatomy and ontogeny. Journal of Vertebrate Paleontology, 26(1), 103–112. doi:10.1671/0272-4634(2006)26[103:TSKTSN]2.0.CO;2 Da extravagante Strukturen aber bei heutigen Tieren nicht unbedingt der Arterkennung dienen, wird diese letzte These heute allerdings auch kontrovers diskutiert.93Hone, D. W. E. & Naish, D. (2013). The ’species recognition hypothesis‘ does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs. Journal of Zoology, 290(3), 172–180. doi:10.1111/jzo.12035

Quellenangaben:
- 1Pearson, D. A. et al. (2002). Vertebrate biostratigraphy of the Hell Creek Formation in southwestern North Dakota and northwestern South Dakota. In Hartman, J. H., Johnson, K. R. & Nichols, D. J. (Hgg.), The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An integrated continental record of the end of the Cretaceous. Geological Society of America, 154. doi:10.1130/0-8137-2361-2.145
- 2Tweet, J. (2025). Paleontology of Morrison–Golden Fossil Areas National Natural Landmark. Park Paleontology News, 17(2).
- 3Marsh, O. C. (1887). Notice of new fossil mammals. American Journal of Science, 34(202), 323–331. doi:10.2475/ajs.s3-34.202.323
- 4Marsh, O. C. (1888). A new family of horned Dinosauria, from the Cretaceous. American Journal of Science, 36(216), 477–478. doi:10.2475/ajs.s3-36.216.477
- 5Marsh, O. C. (1889a). Notice of new American Dinosauria. American Journal of Science, 37(220), 331–336. doi:10.2475/ajs.s3-37.220.331
- 6Marsh, O. C. (1889b). Notice of gigantic horned Dinosauria from the Cretaceous. American Journal of Science, 38(224), 173–175. doi:10.2475/ajs.s3-38.224.173
- 7Cope, E. D. (1872). On the existence of Dinosauria in the Transition Beds of Wyoming. Proceedings of the American Philosophical Society, 12, 481–483.
- 8Cope, E. D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. Bulletin of the U.S. Geological and Geographical Survey of the Territories, 9, 9–28.
- 9Lull, R. S. & Wright, N. E. (1942). Hadrosaurian dinosaurs of North America (Vol. 40). Geological Society of America.
- 10Dodson, P. (1996). The horned dinosaurs. Princeton University Press.
- 11Lull, R. S. (1933). A revision of the Ceratopsia or horned dinosaurs. Memoirs of the Peabody Museum of Natural History, 3(3), 1–175. doi:10.5962/bhl.title.5716
- 12Cope, E.D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. Bulletin of the U.S. Geological and Geographical Survey of the Territories. 9:9-28.
- 13Dodson, P., Forster, C. A. & Sampson, S. D. (2004). Ceratopsidae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Eds.), The dinosauria. University of California Press, 494–513.
- 14Hatcher, J. B., Marsh, O. C. & Lull, R. S. (1907). The Ceratopsia. Government Printing Office.
- 15Mateus, O. (2008). Two ornithischian dinosaurs renamed – Microceratops Bohlin 1953 and Diceratops Lull 1905. Journal of Paleontology, 82(2), 423. doi:10.1666/07-069.1
- 16Scannella, J.; Horner, J.R. (2010). „Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny“. Journal of Vertebrate Paleontology. 30 (4): 1157–1168. Bibcode:2010JVPal..30.1157S. doi:10.1080/02724634.2010.483632. S2CID 86767957.
- 17Farke, A. A. (2011). Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE, 6(1), e16196. doi:10.1371/journal.pone.0016196
- 18Ott, C. J. & Larson, P. L. (2010). A new, small ceratopsian dinosaur from the Latest Cretaceous Hell Creek Formation, northwest South Dakota, United States – a preliminary description. In Ryan, M. J., Chinnery-Allgeier, B. J. & Eberth, D. A. (Eds.), New perspectives on horned dinosaurs – the Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press, 656.
- 19Longrich, N. R. (2011). Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretaceous Research, 32(3), 264–276. doi:10.1016/j.cretres.2010.12.007
- 20Holtz, T. R. Jr. (2011). Dinosaurs – the most complete, up-to-date encyclopedia for dinosaur lovers of all ages, Winter 2010 appendix.
- 21Illies, M. M. et al. (2020). Triceratops with a kink – co-ossification of five distal caudal vertebrae from the Hell Creek Formation of North Dakota. Cretaceous Research, 108, 104355.
- 22Scannella, J. & Horner, J. R. (2010). Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae) – synonymy through ontogeny. Journal of Vertebrate Paleontology, 30(4), 1157–1168. doi:10.1080/02724634.2010.483632
- 23Sullivan, R. M. & Lucas, S. G. (2006). The Kirtlandian land-vertebrate „age“–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America. New Mexico Museum of Natural History and Science Bulletin, 35, 7–29.
- 24Jasinski, S. E., Sullivan, R. M. & Dodson, P. (2020). New dromaeosaurid dinosaur (Theropoda, Dromaeosauridae) from New Mexico and biodiversity of dromaeosaurids at the end of the Cretaceous. Scientific Reports, 10(1), 5105. doi:10.1038/s41598-020-61480-7
- 25Sullivan, R. M. & Lucas, S. G. (2010). A new chasmosaurine (Ceratopsidae, Dinosauria) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. In Ryan, M. J., Chinnery-Allgeier, B. J. & Eberth, D. A. (Eds.), New perspectives on horned dinosaurs – The Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press.
- 26Longrich, N. R. (2011). Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretaceous Research, 32(3), 264–276. doi:10.1016/j.cretres.2010.12.007
- 27Holtz, T. R. Jr. (2011). Dinosaurs – the most complete, up-to-date encyclopedia for dinosaur lovers of all ages, Winter 2010 appendix.
- 28Jasinski, S. E., Sullivan, R. M. & Lucas, S. G. (2011). Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member) San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science Bulletin, 53, 216–271.
- 29Maiorino, L. et al. (2015). Variation in the shape and mechanical performance of the lower jaws in ceratopsid dinosaurs (Ornithischia, Ceratopsia). Journal of Anatomy, 227(5), 631–646. doi:10.1111/joa.12374
- 30Brown, C. M. & Henderson, D. M. (2015). A new horned dinosaur reveals convergent evolution in cranial ornamentation in Ceratopsidae. Current Biology, 25(12), 1641–1648. doi:10.1016/j.cub.2015.04.041
- 31Scannella, J. (2009). And then there was one – synonymy consequences of Triceratops cranial ontogeny. Journal of Vertebrate Paleontology, 29, 177A.
- 32Scannella, J. & Horner, J. R. (2010). Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae) – synonymy through ontogeny. Journal of Vertebrate Paleontology, 30(4), 1157–1168. doi:10.1080/02724634.2010.483632
- 33Farke, A. A. (2011). Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE, 6(1), e16196. doi:10.1371/journal.pone.0016196
- 34Longrich, N. R. & Field, D. J. (2012). Torosaurus is not Triceratops – ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy. PLoS ONE, 7(2), e32623. doi:10.1371/journal.pone.0032623
- 35Mallon, J. C. et al. (2022). The record of Torosaurus (Ornithischia: Ceratopsidae) in Canada and its taxonomic implications. Zoological Journal of the Linnean Society, 195(1), 157–171. doi:10.1093/zoolinnean/zlab120
- 36Scannella, J. B. & Fowler, D. W. (2009). Anagenesis in Triceratops – evidence from a newly resolved stratigraphic framework for the Hell Creek Formation. 9th North American Paleontological Convention Abstracts. Cincinnati Museum Center Scientific Contributions, 3, 148–149.
- 37Mallon, J. C. et al. (2025). The Canadian fossil record supports anagenesis in Triceratops (Ornithischia, Ceratopsia). Canadian Journal of Earth Sciences, 62(7), 1222–1236. doi:10.1139/cjes-2024-0170
- 38Lambe, L. M. (1915). On Eoceratops canadensis, gen. nov., with remarks on other genera of Cretaceous horned dinosaurs. Museum Bulletin No. 12, Geological Series No. 24. Geological Survey, Department of Mines, Ottawa.
- 39Madzia, D. et al. (2021). The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ, 9, e12362. doi:10.7717/peerj.12362
- 40Hatcher, J. B., Marsh, O. C. & Lull, R. S. (1907). The Ceratopsia. Government Printing Office, Washington, D.C.
- 41Lull, R. S. (1933). A revision of the Ceratopsia or horned dinosaurs. Memoirs of the Peabody Museum of Natural History, 3(3), 1–175. doi:10.5962/bhl.title.5716
- 42Sternberg, C. M. (1949). The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member. Phylogeny of the Ceratopsidae. National Museum of Canada Bulletin, 113, 33–46.
- 43Ostrom, J. H. (1966). Functional morphology and evolution of the ceratopsian dinosaurs. Evolution, 20(3), 290–308. doi:10.2307/2406631
- 44Norman, D. (1985). The Illustrated Encyclopaedia of Dinosaurs. Salamander Books, London.
- 45Granger, W. W. & Gregory, W. K. (1923). Protoceratops andrewsi, a pre-ceratopsian dinosaur from Mongolia. American Museum of Natural History Novitates, 72, 1–9.
- 46Wolfe, D. G. & Kirkland, J. I. (1998). Zuniceratops christopheri n. gen. & n. sp., a ceratopsian dinosaur from the Moreno Hill Formation (Cretaceous, Turonian) of west-central New Mexico. New Mexico Museum of Natural History and Science, Bulletin, 14, 303–317.
- 47Xu, X., Forster, C. A., Clark, J. M. & Mo, J. (2006). A basal ceratopsian with transitional features from the Late Jurassic of northwestern China. Proceedings of the Royal Society B: Biological Sciences, 273(1598), 2135–2140. doi:10.1098/rspb.2006.3566
- 48Gauthier, J. A. (1986). Saurischian monophyly and the origin of birds. The Origin of Birds and the Evolution of Flight, K. Padian (Hg.). Memoirs of the California Academy of Sciences, 8, 1–55.
- 49Sereno, P. C. (1998). A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 210(1), 41–83. doi:10.1127/njgpa/210/1998/41
- 50Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press, 265–267.
- 51Lambert, D. (1993). The Ultimate Dinosaur Book. Dorling Kindersley, New York. pp. 152–167. ISBN 978-1-56458-304-8.
- 52Tait, J. & Brown, B. (1928). How the Ceratopsia carried and used their head. Transactions of the Royal Society of Canada, 22, 13–23.
- 53Ostrom, J. H. (1966). Functional morphology and evolution of the ceratopsian dinosaurs. Evolution, 20(3), 290–308. doi:10.2307/2406631
- 54Erickson, G. M. et al. (2015). Wear biomechanics in the slicing dentition of the giant horned dinosaur Triceratops. Science Advances, 1(5), e1500055. doi:10.1126/sciadv.1500055
- 55Maiorino, L., Farke, A. A., Kotsakis, T., Teresi, L. & Piras, P. (2015). Variation in the shape and mechanical performance of the lower jaws in ceratopsid dinosaurs (Ornithischia, Ceratopsia). Journal of Anatomy, 227(5), 631–646. doi:10.1111/joa.12374
- 56Coe, M. J., Dilcher, D. L., Farlow, J. O., Jarzen, D. M. & Russell, D. A. (1987). Dinosaurs and land plants. In Friis, E. M., Chaloner, W. G. & Crane, P. R. (Hrsg.), The Origins of Angiosperms and their Biological Consequences. Cambridge University Press, 225–258.
- 57de Rooij, J. et al. (2022). Stable isotope record of Triceratops from a mass accumulation (Lance Formation, Wyoming, USA) provides insights into Triceratops behaviour and ecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 607, 111274. doi:10.1016/j.palaeo.2022.111274
- 58Fricke, H. C. & Pearson, D. A. (2008). Stable isotope evidence for changes in dietary niche partitioning among hadrosaurian and ceratopsian dinosaurs of the Hell Creek Formation, North Dakota. Paleobiology, 34(4), 534–552. doi:10.1666/08020.1
- 59Sander, P. M., Gee, C. T., Hummel, J. & Clauss, M. (2010). Mesozoic plants and dinosaur herbivory. In Gee, C. T. (Hg.), Life of the Past, 331–359. doi:10.5167/uzh-35283
- 60Barrett, P. M. & Rayfield, E. J. (2006). Ecological and evolutionary implications of dinosaur feeding behaviour. Trends in Ecology & Evolution, 21(4), 217–224. doi:10.1016/j.tree.2006.01.002
- 61Dodson, P., Forster, C. A. & Sampson, S. D. (2004). Ceratopsidae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Hrsg.), The Dinosauria. University of California Press, Berkeley, 494–513.
- 62Scannella, J. B. et al. (2014). Evolutionary trends in Triceratops from the Hell Creek Formation, Montana. Proceedings of the National Academy of Sciences, 111(28), 10245–10250. doi:10.1073/pnas.1313334111
- 63Christiansen, P. & Paul, G. S. (2001). Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs. Gaia, 16, 13–29.
- 64Thompson, S. & Holmes, R. (2007). Forelimb stance and step cycle in Chasmosaurus irvinensis (Dinosauria: Neoceratopsia). Palaeontologia Electronica, 10(1), 17.
- 65Fujiwara, S.-I. (2009). A reevaluation of the manus structure in Triceratops (Ceratopsia: Ceratopsidae). Journal of Vertebrate Paleontology, 29(4), 1136–1147. doi:10.1671/039.029.0406
- 66Rega, E., Holmes, R. & Tirabasso, A. (2010). Habitual locomotor behavior inferred from manual pathology in two Late Cretaceous chasmosaurine ceratopsid dinosaurs, Chasmosaurus irvinensis (CMN 41357) and Chasmosaurus belli (ROM 843). In Ryan, M. J., Chinnery-Allgeier, B. J. & Eberth, D. A. (Hgg.), New Perspectives on Horned Dinosaurs. The Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press, Bloomington and Indianapolis, 340–354.
- 67Mathews, J. C., Brusatte, S. L., Williams, S. A. & Henderson, M. D. (2009). The first Triceratops bonebed and its implications for gregarious behavior. Journal of Vertebrate Paleontology, 29(1), 286–290. doi:10.1080/02724634.2009.10010382
- 68Smith, M. (2013). Triceratops trio unearthed in Wyoming. CNN.
- 69Illies, M. M. C. & Fowler, D. W. (2020). Triceratops with a kink. Co-ossification of five distal caudal vertebrae from the Hell Creek Formation of North Dakota. Cretaceous Research, 108, 104355. doi:10.1016/j.cretres.2019.104355
- 70Fowler, Denver & Scannella, John & Goodwin, Mark. (2012). How to eat a Triceratops: large sample of toothmarks provides new insight into the feeding behavior of Tyrannosaurus. J. Vert. Paleontol.. 32. 96-96.
- 71Sakagami, R. & Kawabe, S. (2020). Endocranial anatomy of the ceratopsid dinosaur Triceratops and interpretations of sensory and motor function. PeerJ, 8, e9888. doi:10.7717/peerj.9888
- 72Hu, J., Xu, X., Zhao, Q., He, Y., Forster, C. A. & Han, F. (2024). Endocranial morphology of three early-diverging ceratopsians and implications for the behavior and the evolution of the endocast in ceratopsians. Paleobiology, 50(3), 490–502. doi:10.1017/pab.2024.25
- 73Tada, S. et al. (2026). Nasal soft-tissue anatomy of Triceratops and other horned dinosaurs. The Anatomical Record. doi:10.1002/ar.70150
- 74de Rooij, J. et al. (2022). Stable isotope record of Triceratops from a mass accumulation (Lance Formation, Wyoming, USA) provides insights into Triceratops behaviour and ecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 607, 111274. doi:10.1016/j.palaeo.2022.111274
- 75Sereno, P. C. et al. (2026). Duck-billed dinosaur fleshy midline and hooves reveal terrestrial clay-template „mummification“. Science, 391(6780), eadw3536. doi:10.1126/science.adw3536
- 76Bell, P. R. et al. (2022). The exquisitely preserved integument of Psittacosaurus and the scaly skin of ceratopsian dinosaurs. Communications Biology, 5(1), 809. doi:10.1038/s42003-022-03749-3
- 77Sternberg, C. H. (1917). Hunting dinosaurs in the Badlands of the Red Deer River, Alberta, Canada. C. H. Sternberg—The World Company Press. doi:10.5962/bhl.title.159811
- 78Bakker, R. T. (1986). The dinosaur heresies – New theories unlocking the mystery of the dinosaurs and their extinction. William Morrow, 438
- 79Happ, J. (2008). An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. In Larson, P. & Carpenter, K. (Eds.), Tyrannosaurus rex, the Tyrant King (Life of the Past). Indiana University Press, 355–368.
- 80Erickson, G. M. & Olson, K. H. (1996). Bite marks attributable to Tyrannosaurus rex – preliminary description and implications. Journal of Vertebrate Paleontology, 16(1), 175–178. doi:10.1080/02724634.1996.10011297
- 81Farke, A. A. et al. (2009). Evidence of combat in Triceratops. PLoS ONE, 4(1), e4252. doi:10.1371/journal.pone.0004252
- 82Reid, R. E. H. (1997). Histology of bones and teeth. In Currie, P. J. & Padian, K. (Eds.), Encyclopedia of dinosaurs. Academic Press, 329–339.
- 83Horner, J. R. & Lamm, E. (2011). Ontogeny of the parietal frill of Triceratops – a preliminary histological analysis. Comptes Rendus Palevol, 10(5–6), 439–452. doi:10.1016/j.crpv.2011.04.006
- 84Farlow, J. O. & Dodson, P. (1975). The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. Evolution, 29(2), 353–361. doi:10.2307/2407222
- 85D’Anastasio, R. et al. (2022). Histological and chemical diagnosis of a combat lesion in Triceratops. Scientific Reports, 12(1), 3941. doi:10.1038/s41598-022-08033-2
- 86Wheeler, P. E. (1978). Elaborate CNS cooling structures in large dinosaurs. Nature, 275(5679), 441–443. doi:10.1038/275441a0
- 87Davitashvili, L. Sh. (1961). Teoriya Polovogo Otbora (Theory of Sexual Selection). Izdatel’stvo Akademii nauk SSSR, 538.
- 88Farlow, J. O. & Dodson, P. (1975). The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. Evolution, 29(2), 353–361. doi:10.2307/2407222
- 89Lehman, T. M. (1990). The ceratopsian subfamily Chasmosaurinae – sexual dimorphism and systematics. In Carpenter, K. & Currie, P. J. (Eds.), Dinosaur Systematics – Perspectives and Approaches. Cambridge University Press, 211–229.
- 90Forster, C. A. (1990). The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny. Ph.D. Dissertation thesis, University of Pennsylvania, 227.
- 91Dodson, P., Forster, C. A. & Sampson, S. D. (2004). Ceratopsidae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (Eds.), The dinosauria. University of California Press, 494–513.
- 92Goodwin, M. B. et al. (2006). The smallest known Triceratops skull – new observations on ceratopsid cranial anatomy and ontogeny. Journal of Vertebrate Paleontology, 26(1), 103–112. doi:10.1671/0272-4634(2006)26[103:TSKTSN]2.0.CO;2
- 93Hone, D. W. E. & Naish, D. (2013). The ’species recognition hypothesis‘ does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs. Journal of Zoology, 290(3), 172–180. doi:10.1111/jzo.12035
Hat dir dieser Beitrag gefallen?
Für noch mehr Lesestoff schau gerne in die Kategorie Meine Bücher. Alle meine Werke findest du auch in der Sidebar rechts. Dort gibt es auch einen Direktlink zu Amazon, wo du sie gleich bestellen kannst. ➡️
Wie kannst du Die Weißen Steine sonst noch unterstützen?
Klar, in erster Linie natürlich, wenn du dir eines meiner Bücher kaufst. Aber du kannst mir auch auf anderen Wegen eine große Freude machen:
- Teile diesen Beitrag auf deinen Social-Media-Kanälen!
- Schreibe mir eine nette Rezension auf Amazon!
- Kaufe hier auf der Website über Affiliate Links ein!
Ich kriege übrigens auch eine kleine Unterstützung, wenn du etwas völlig anderes kaufst. Es muss nicht unbedingt der hier beworbene Artikel sein! Jeder kleine Centbetrag hilft, diese Seite am Laufen zu erhalten.
 |
Zusammen mit meiner lieben Frau Sahar habe ich auch noch eine Menge mehr zu bieten! Sahar ist nämlich eine großartige Künstlerin und fertigt Gemälde, Lesezeichen, Stofftaschen und viele andere coole Artikel an. Auch mit tollen Urzeit-Motiven! Klick dich einfach mal durch die Beiträge in der Kategorie Produkte & Services. Bestimmt findest du da etwas nach deinem Geschmack. Oder auch ein schönes Geschenk für deine Lieben. |
Einen Grund, anderen eine Freude zu bereiten, findet man schließlich immer!
Thanks for sharing. I read many of your blog posts, cool, your blog is very good.